pigmentation cutanée/pigmentation
Chez l’Homme, la couleur de la peau et des poils est principalement déterminée par le nombre, la taille, le type et le mode de répartition des mélanosomes . Il est particulièrement intéressant de noter que dans les conditions normales, les différences raciales de pigmentation de la peau chez l’Homme ne reposent pas sur des différences numériques de la population mélanocytaire épidermique. Pour une zone déterminée, le nombre de mélanocytes épidermiques est sensiblement identique chez le noir, le blanc ou l’asiatique. Les facteurs prépondérants dans le déterminisme de la couleur de la peau sont donc le type de pigment synthétisé et le niveau d’activité des mélanocytes.
Les principales étapes qui déterminent la pigmentation constitutive de la peau et qui sont les mêmes dans les peaux de différents groupes ethniques ou les différentes races, sont : la migration des mélanoblastes vers l’épiderme , leur survie et leur différenciation en mélanocytes, la densité des mélanocytes, l’expression et la fonction des constituants enzymatiques et structuraux des mélanosomes, la synthèse des différents types de mélanine (eu- et pheomélanine), le transport des mélanosomes aux dendrites du mélanocyte, le transfert des mélanosomes aux kératinocytes et finalement la distribution des mélanines et leur dégradation au niveau de la peau.
La pigmentation peut être modulée par un grand nombre de facteurs intrinséques ou extrinséques, tels que la région du corps, les différences de sexe, d’ethnies, les anomalies génétiques, l’âge, des réponses variables aux signaux hormonaux, des changements liés au cycle pilaire, le climat, les saisons, l’exposition aux UV , le contact avec des toxines, des polluants, des infections microbiennes.
Des facteurs intrinséques qui régulent la pigmentation peuvent provenir non seulement des kératinocytes et des fibroblastes mais également des cellules endothèliales et des hormones véhiculées par l’apport sanguin, des cellules inflammatoires et du système nerveux.
Dans les keratinocytes, Foxn1 et p53 sont des facteurs de transcription qui régulent la pigmentation cutanée via le FGFbéta et des dérivés de POMC tels que la α-MSH et l’ACTH, respectivement. D’autres activateurs kératinocytaires de la mélanogénèse sont le SCF/steel factor (stem cell factor ), l’HGF (hepatocyte growth factor ), le GM-CSF (granulocyte-macrophage colony-stimulating factor), le NGF (nerve growth factor ), l’endorphine, l’endotheline-1 (ET-1), la prostaglandine (PG)E2/PGF2α et le LIF( leukemia inhibitory factor ).
Dickkopf 1 (DKK1) est un inhibiteur de la voie de signalisation Wnt qui est abondamment secrété dans le derme des plantes des pieds et des paumes des mains. DKK1 inhibe la prolifération et la fonction des mélanocytes en inhibant la voie de la mélanogénèse au niveau de la transcription des facteurs comme Mitf et de la production des protéines mélanogéniques. DKK1 affecte également le transfert de la mélanine des mélanocytes aux kératinocytes en supprimant l’expression de PAR-2.
Les cellules endothéliales sont des sources d’endothéline-1, de prostaglandines PGE2/PGF2, et d’oxyde nitrique (NO) qui active la pigmentation cutanée.
Les nerfs produisent différents stimulateurs des mélanocytes, tels que le NGF (Nerve Growth Factor) ou la CGRP (calcitonin gene- related peptide).
Les estrogènes stimulent la pigmentation et les androgènes l’inhibent. Les produits de clivage de la pro-opiomelanocortin, alpha-MSH, ACTH, et endorphine sont des facteurs hormonaux qui stimulent la mélanogénèse.
Les prostaglandines, les thromboxanes, et les leucotriènes augmentent l’activité tyrosinase et sont responsables de l’hyperpigmentation post inflammatoire. Par contre, l’IL6 et le TNFalpha sont des inhibiteurs de la pigmentation cutanée. L’histamine , le NO, le GM-CSF, et l’alpha-MSH sont d’autres facteurs produits au cours de l’inflammation qui augmentent la mélanogénèse.
La pigmentation induite par les UVs
L’irradiation UV est également bien connue pour augmenter la plupart des facteurs qui stimulent la mélanogénèse. Les UVs induisent une réponse immédiate et une réponse plus tardive. L’ action immédiate (quelques minutes) persiste plusieurs jours mais cette augmentation rapide de la pigmentation résulte seulement de l’oxydation de pigments préexistants et de la redistribution des mélanosomes sans augmentation de la mélanogenèse. La réponse tardive aux UVs correspond à une augmentation de la mélanogénèse qui résulte d’ une augmentation de l’expression de MITF, un régulateur majeur de la transcription de la pigmentation et de ces cibles en aval incluant Pmel17, MART-1, la tyrosinase, Tyrp1, Tyrp2 / Dct. De plus, les mélanocytes épidermiques et également les kératinocytes répondent à une exposition aux UVs en augmentant leurs productions en alpha-MSH et ACTH, qui, à leur tour, induisent une augmentation de l’expression de MC1R à la surface des mélanocytes et stimulent ainsi la mélanogénèse.
Les personnes à peau claire, avec une faible habilité à bronzer, ou présentant des tâches de rousseur avec ou sans cheveux roux, ont un risque double d’avoir un mélanome. Les polymorphismes du gène MC1R sont associés à ce risque. Le mélanome est rarement présent chez les personnes n’ayant pas la peau blanche, avec une incidence 10 à 20 fois moins élevée.
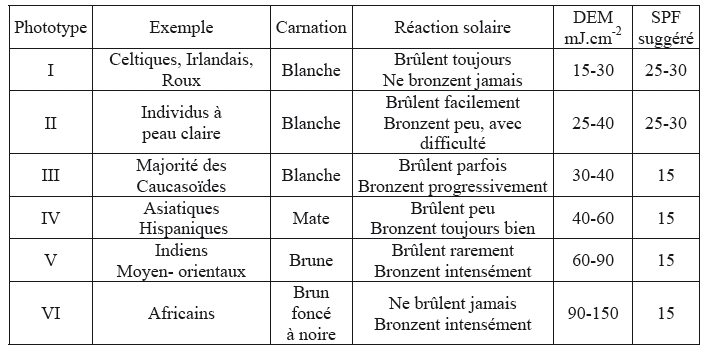
- Phototypes, DEM et SPF
Articles
Le vieillissement cutané
Effets tissulaires des UV
Immunosuppression induite par les UVs
Le mélanocyte et la pigmentation
Les produits dépigmentants
L’EPIDERME, UNE BARRIERE SUR TOUS LES FRONTS
Mots-clés dans le même groupe
Autres groupes de mots-clés
Rubriques
Site réalisé avec SPIP 3.0.17 + AHUNTSIC

Visiteurs connectés : 46