Post a message
Replying to:
Melanocyte and pigmentation
Normal human skin color ranges from white to brown to black and results from the mixed contribution of four pigments: oxygenated hemoglobin (red) in the capillaries, reduced hemoglobin (blue) in the venules of the dermis, exogenously produced carotenoids (yellow) and endogenously produced melanin (brown). However, it is the amount and type of melanin pigment produced in melanosomes by cutaneous and follicular melanocytes which largely determined the differences in skin and hair color between individuals.
Normal skin pigmentation is a complex process that, in the epidermis as in the hair follicles, begins with the synthesis of melanin within melanosomes in the melanocytes, followed by melanosome transfer to neighboring basal and suprabasal keratinocytes. In basal cells, melanin granules are translocated to the upper pole of the nucleus, forming a melanin cap that protect DNA from UV rays. Melanin granules are eventually degraded as the keratinocyte undergoes terminal differentiation.
1. Melanocytes
Melanocytes are specialized cells of the epidermis that produce the pigment melanin. There are approximately 2000 or more melanocytes per square millimeter in the exposed skin of the head, in the skin of the scrotum or in the foreskin and 1,000 to 1,500 melanocytes per square millimeter on the rest of the body in Caucasoids, Negroids and Mongoloids. Accordingly, racial differences in pigmentation are not due to a difference in the number of melanocytes.
Epidermal melanocytes are regularly dispersed, at an approximate ratio of 1:10, among basal keratinocytes and distribute the melanin they produce to around 40 overlying suprabasal keratinocytes via their elongated dendrites and cell/cell contacts. The anatomical relationship between keratinocytes and melanocytes is known as “the epidermal melanin unit”. The secretory melanocytes in hair follicles are associated with cells of the hair matrix in the same way that the epidermal melanocytes are associated with the epidermal keratinocytes.
The epidermal melanocyte activity is continuous while the melanocytes of the hair follicle follow its rhytmical activity; they are active during the anagen phase of growth and have no detectable tyrosinase activity during the resting (telogen) phase.
Melanocyte proliferation occurs in non stimulated, unexposed human skin and is increased after UV irradiation. In hair follicle, melanocyte proliferation and melanin synthesis are both synchronized with hair growth.
Melanocytes are detected in histological sections by ammoniacal silver nitrate reaction of Fontana-Masson which stains melanins and by DOPA reaction which revealed tyrosinase activity. They are stained in immunohistology with various antibodies against melanocyte antigens, such as tyrosinase, pigmentation associated protein and MART1/Melan-A antigen. Embryonic melanocytes, hair bulb melanocytes and activated (but not resting) melanocytes of adult epidermis express the premelanosome-associated glycoprotein gp100 recognised by the monoclonal antibody HMB-45.
Ultrastructurally, they have an electron-lucent cytoplasm containing no tonofilaments and no desmosomes but containing loose intermediate filaments (vimentin) and specific organelles, the melanosomes at various maturation stages.
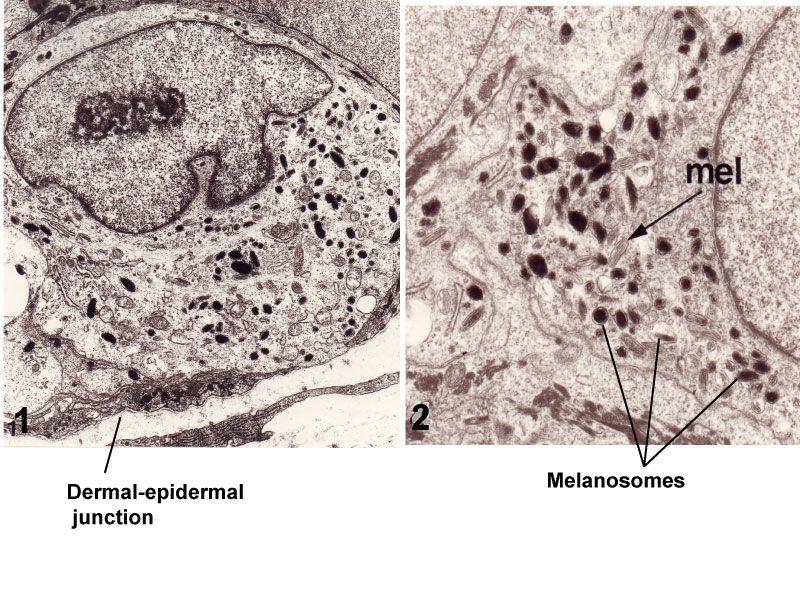
- Human melanocyte observed by transmission electron microscopy in a human skin transplanted onto the nude mouse (x10000); 2) higher magnification of an other human melanocyte showing melanosomes (x 20000).
- Note the presence of melanosomes at various stages of differentiation, the absence of desmosomes with neighbouring keratinocytes and the contact with the dermal epidermal junction without lamina densa.
The melanocytes are derived from precursor cells (called melanoblasts) during embryological development, and melanoblasts destined for the skin originate from the neural crest beginning in the second month of human embryonic life. The melanoblasts are large round or oval cells which differentiate into melanocytes by becoming dendritic and DOPA oxydase-positive. They reach dermis between the 10th and the 12th week of development and 2 weeks later for the epidermis and differentiate into melanocytes; melanocytes are already established at epidermal-dermal junction sites at about the sixth month of fetal life. The melanocytes colonise the dermis and the epidermis before the initial stage of hair production and distribute into the hair without a specific localisation. It is only after the sixth month of gestation, that, in the hair, the melanocytes will be strictly localised in the infundibulum and in the hair matrix. Dermal melanocytes decrease in number during gestation and virtually disappear by birth, whereas epidermal melanocytes established at the epidermal-dermal junction continue to proliferate and start to produce melanosomes in which melanin is synthesized.
2. Melanosomes
Melanosomes are specific melanin-producing intracellular organelles that share several features with lysosomes in that they contain acid dependent hydrolases and lysosomal-associated membrane proteins (LAMPs). They belong to a family of cell-specific organelles, termed lysosome-related organelles , which also include lytic granules observed in cytotoxic T lymphocytes and natural killer cells, MHC class II compartments observed in antigen presenting cells, platelet-dense granules, basophil granules, azurophil granules observed in neutrophils and Weibel-Palade bodies observed in endothelial cells. The main structural component of melanosomes is Pmel17/gp100/Silv.
Melanosomes result from the fusion of vesicles that contain tyrosinase (Tyr), dopachrome tautomerase (Dct/Tyrp2) and DHCI oxydase (Tyrp1) and derive from Golgi apparatus with vesicles that contain the structural components of melanosomes and originate from granular endoplasmic reticulum. The trafficking of Tyr to melanosomes can be affected dramatically by mutations in several melanogenic genes and by changes of intracellular pH that result in oculocutaneous albinism or hypopigmentation, respectively.
2.1 Structure of melanosomes
The melanosome structure is different according to the type of melanin produced; pheomelanosomes produce pheomelanin, remain round and contain vesiculoglobular matrix; eumelanosomes produce eumelanin, are elliptical and have a fibrillar matrix.
Both types of melanosomes are typically divided into four maturation stages (I–IV) determined by their structure and the quantity, quality, and arrangement of the melanin produced. Stage I melanosomes have an early matrix organization, are spherical, do not contain tyrosinase activity, and are localised at the periphery of the nucleus. Stage II melanosomes are tyrosinase-containing elongated (for eumelanosomes) organelles with an organized filamentous matrix and with no melanin deposition in eumelanosomes and already formed melanin and a vesiculo-globular matrix in phaeomelanosomes.The production of the internal matrix fibers and the maturation from stage I to stage II appear dependent of the presence of a structural protein named Pmel17 (or gp 100/Silver). Just after its transfer into the stage I melanosome, Pmel17 is cleaved into several fragments which constitute the fibrillar matrix of the organite. Another protein, Mart-1, also known as Melan-A, localised in stage I and/or II, contributes to the melanosome formation since it is necessary to the expression, the stability and the maturation of Pmel17 and therefore to the critical step of the stage II melanosome formation.
In type III melanosomes, melanin is uniformly deposited. Type IV melanosomes are melanosomes that are electron-opaque, fully melanized and have a low tyrosinase activity; they are the melanosomes supplied from the dendrites to the neighboring keratinocytes.
2.2 The enzymes of pigmentation
Within melanosomes, at least three key-enzymes, named tyrosinase, Tyrosinase-related protein 1 (Tyrp1), and Tyrosinase-related protein 2 (Tyrp2/Dct) are absolutely required for the synthesis of different types of melanin.
Tyrosinase is responsible for the critical initial rate limiting steps of melanogenesis. It is synthetised as an inactive precursor that is activated when melanocytes are stimulated by alpha-MSH via AMPc. The Agouti protein is an alpha-MSH antagonist. It blocks the alpha-MSH fixation to its receptor and, therefore, the receptor activation and stabilises the inactive form of the receptor. Tyrosinase is dependent upon the incorporation of copper ion to catalyze the hydroxylation of tyrosine to β-3,4-dihydroxyphenylalanine (DOPA) and the subsequent oxidation of DOPA to DOPAquinone. It is also involved downstream in the oxydation of 5,6-dihydroxyindole (DHI) to indole-5,6-quinone.
Tyrp1 and Tyrp2/Dct have distinct catalytic functions in melanin synthesis downstream of Tyr. The isomerisation of DOPAchrome (indolene-2-carboxylic acid-5,6-quinone) to 5,6-dihydroxyindole-2-carboxylic acid (DHICA) is catalysed by DOPAchrome tautomerase (Tyrp2/Dct) and the oxydation of DHICA is catalysed by DHICA-oxydase (Tyrp1). In addition to having enzymatic functions, Tyrp1 and Tyrp2/Dct also stabilize Tyr. Tyrp1 is also a critical enzyme for the correct trafficking of tyrosinase to melanosomes and Tyrp2/Dct seems to also play an important role in detoxification processes within melanosomes.
2.3 Transport and transfer of melanosomes
During their maturation, melanosomes move from the perinuclear area of melanocytes where they are produced towards the extremity of dendrites as they become more melanized. This microtubule-dependent intracellular transport involves microtubules, dynein and kinesin, actin filaments, Rab27a, melanophilin, myosin Va and Slp2-a. Melanin synthesized in melanosomes within melanocytes is transferred to keratinocytes through the protease-activated receptor, PAR-2.
3. Melanins
Mammalian melanocytes produce two chemically distinct types of melanin pigments: black-brown eumelanin and yellow-reddish pheomelanin .
Eumelanin is a highly heterogeneous polymer consisting of DHI (DiHydroxyIndole) and DHICA ((DiHydroxyIndole Carboxylic Acid) units in reduced or oxidized states. Pheomelanin consists mainly of sulfur-containing benzothiazine derivatives. Melanins are synthesized from tyrosine exogenously supplied by blood. Tyrosines are oxidized by tyrosinase and metabolised into DOPAs and then into DOPAquinones which are automatically oxidized into indole compounds. Indole compounds connect to each other to produce eumelanins.
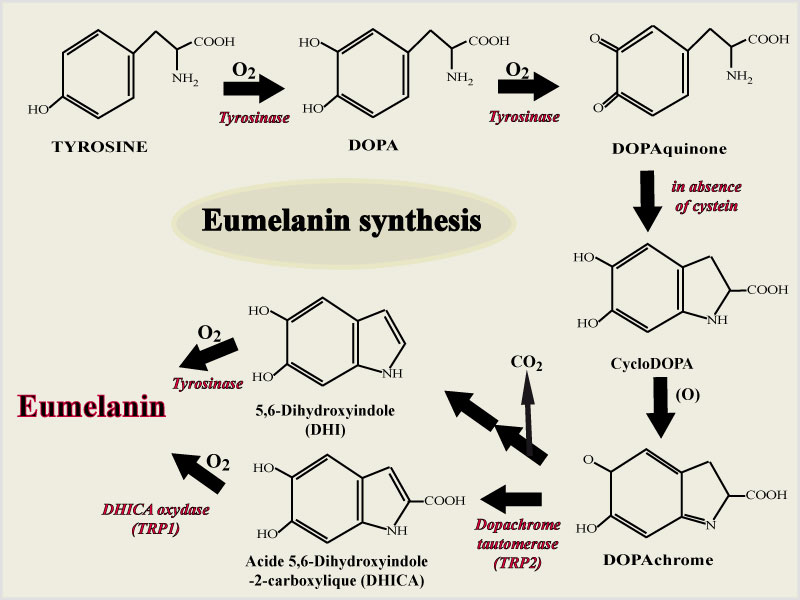
- The different steps of eumelanin synthesis.
The pheomelanin synthesis pathway involves sulfur compounds, the amino acid cystein or glutathion that liberates cysteins through the action of a glutamyl-transpeptidase. In presence of cysteins, DOPAquinones connect with cysteins to form 5-S-cysteinyl DOPA and 2-S-cysteinylDOPA which give benzothiazin intermediates that polymerise to produce pheomelanins.
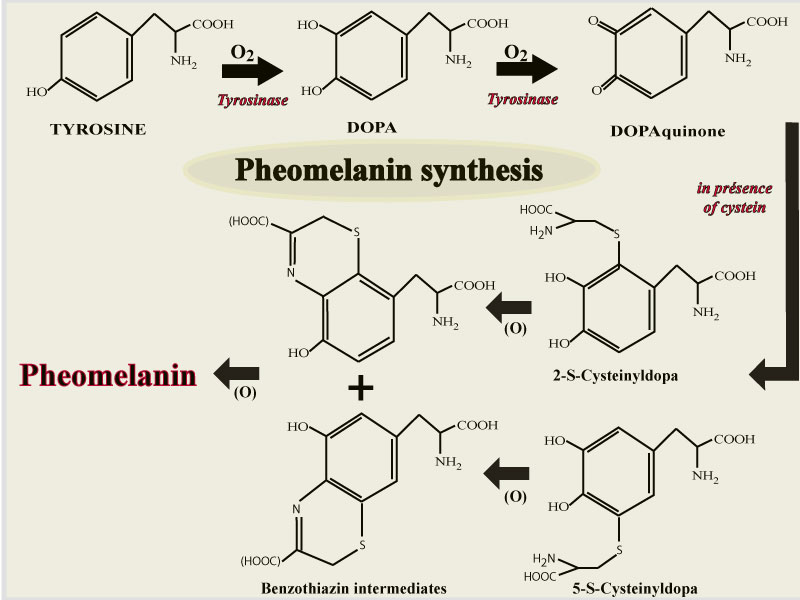
The major role of melanins is to protect skin from the harmful effects of UV rays and to prevent skin cancer . Besides this, both eumelanin and pheomelanin play an important protective role within melanocytes and keratinocytes due to their ability to bind cations, anions, drugs, and chemicals.
4. Cutaneous pigmentation
In human, as in the others mammals, the skin and hair color is mainly determined by the number, the size, the type, and the mode of repartition of the melanosomes. It is particularly interesting to note that in normal conditions, the racial differences in skin pigmentation in human do not depend of the melanocyte number in the epidermis. For a specific area, the number of epidermal melanocytes is nearly the same in Caucasoids, Negroids and Mongoloids, the number, size, and manner in which melanosomes are distributed within keratinocytes vary. The determining factors in skin color are the type of melanin and the level of melanocyte activity. There are differences in melanin content of human melanocytes not only between different skin types but also between different sites of the skin from the same individual.
Pigmentation is classically distinguished into constitutive pigmentation or facultative pigmentation. Constitutive skin pigmentation corresponds to a genetically determined level of cutaneous melanin, in the absence of acquired exogenous or endogenous influences. Facultative pigmentation, on the other hand, designates an induced level of increased epidermal melanin content as a result of solar radiation , hormones or other environmental factors.
The mechanisms involved in constitutive skin pigmentation are similar in skin of different racial/ethnic groups. As previously said, cutaneous pigmentation results from two major events: the synthesis of melanin by melanosomes in melanocytes and the transfer of melanosomes to surrounding keratinocytes; it depends on several factors including: the melanocyte number, the expression and function of melanosomal enzymes and structural proteins, the amount of eu- and pheomelanin synthesized, the transport of melanosomes to dendrites, their transfer to keratinocytes and the distribution of melanin in suprabasal epidermal layers, and eventually, the degradation of melanins in the differentiated keratinocytes.
4.1 Factors that regulate skin pigmentation
A large number of intrinsic and extrinsic factors, such as the body distribution, ethnicity/gender differences, genetic defects, age, variable hormone-responsiveness, hair cycle-dependent changes, climate/season, UV-radiation, toxin, pollutants, chemical exposure and infestations affect skin pigmentation.
Intrinsic factors that regulate pigmentation originate not only from keratinocytes and fibroblasts but also from endothelial cells and hormones via the blood supply, from inflammatory cells and from the nervous system.
In keratinocytes, Foxn1 and p53 are transcription factors that up-regulate skin pigmentation via bFGF and POMC derivatives including α-MSH and ACTH, respectively. Other keratinocyte-derived activators of melanogenesis are stem cell factor (SCF/steel factor), hepatocyte growth factor (HGF), granulocyte-macrophage colony-stimulating factor (GM-CSF), nerve growth factor (NGF), endorphin, endothelin-1 (ET-1), prostaglandin (PG)E2/PGF2α and leukemia inhibitory factor (LIF).
Dickkopf 1 (DKK1) is an inhibitor of the Wnt signaling pathway and a head inducer during development, which is abundantly expressed in palmoplantar fibroblasts and is responsible for thickened and hypopigmented palmoplantar epidermis. DKK1 not only affects melanocytes and suppresses their proliferation and differentiation, but also decreases melanin transfer from melanocytes to keratinocytes via the suppression of PAR-2, explaining why palmoplantar epidermis is hypopigmented. Transforming growth factor-β1 (TGFβ1) is an other fibroblast-derived inhibitor of melanogenesis. Other fibroblast-derived activators of melanocytes include bFGF, HGF and SCF which are also secreted from keratinocytes.
Endothelial cells are sources of endothelin-1, prostaglandin (PG)E2/PGF2α and nitric oxide (NO) which upregulate skin pigmentation.
Nerves produces various melanocyte stimulators such as nerve growth factor (NGF) or calcitonin gene-related peptide (CGRP ).
Estrogens stimulate pigmentation and androgen have inhibitory effects. The products of cleavage of pro-opiomelanocortin ,α-MSH, ACTH and endorphin, are other hormonal factors that stimulate melanogenesis.
Prostaglandins, thromboxanes and leukotrienes are tyrosinase enhancers and are responsible for the hyperpigmentation observed during inflammation . On the other hand, interleukins IL-1, IL-6 and tumor necrosis factor-α (TNFα) are suppressors of skin
pigmentation. Histamine , NO, GM-CSF and α-MSH are other factors produced during inflammation that increase melanogenesis.
4.2 UV-induced pigmentation
UV radiation is well known to up-regulates most factors that increase melanogenesis and to increase skin pigmentation.
UV, especially UVA, induce an immediate skin darkening that occurs within minutes and persists for several hours followed by persistent pigment darkening, which occurs for several hours and lasts for several weeks. This rapid increase in pigmentation result from the oxidation and polymerization of existing melanins and of the redistribution of existing melanosomes; it does not result from an acute melanin synthesis.
Delayed UV response corresponds to an increase in melanogenesis which results from an increase in expression of MITF, a major regulator of pigmentation transcription pathway and of its downstream targets, including Pmel17, MART-1, tyrosinase, Tyrp1, Tyrp2/DCT. Moreover, the epidermal melanocytes and also the keratinocytes, respond to UV radiation by increasing alpha-MSH and ACTH productions which will induce an increase in MC1R at the melanocyte surface and thus stimulate melanogenesis.
PAR-2 is also increased by UV exposure in keratinocytes and induces an increased uptake of melanosomes by keratinocytes.
4.3 Fitzpatrick’s Skin Phototypes
Site powered by SPIP 3.0.17 + AHUNTSIC

Visitors logged in: 77