Post a message
Replying to:
The stem cells
A stem cell is a cell found in all multicellular organisms that possesses two properties:
- it can self-renew to produce more stem cell
- it can differentiate into diverse specialized cell types.
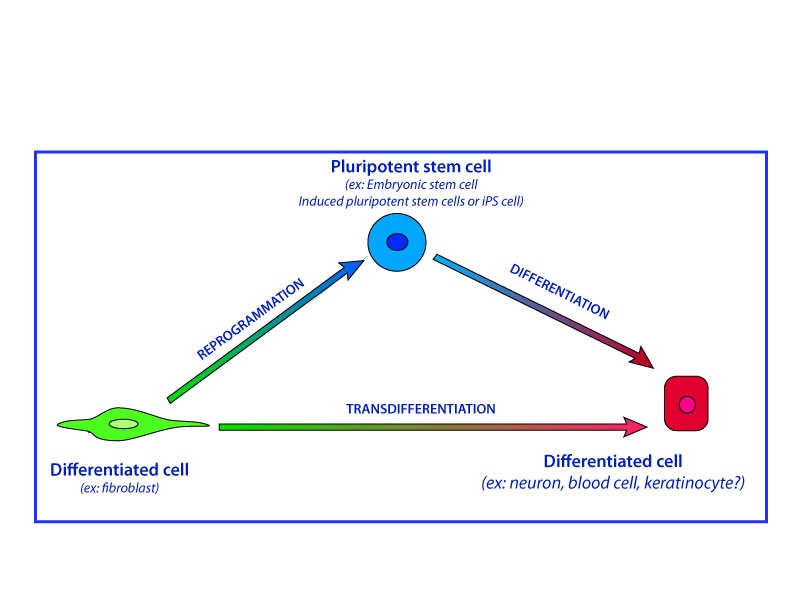
- The different pathways of transformation from a cell type to another one. (adapted from the blog of Albert Barrois)
1. Differentiation potency
Several types of stem cells are distinguished according to the number of different cell types they are able to generate:
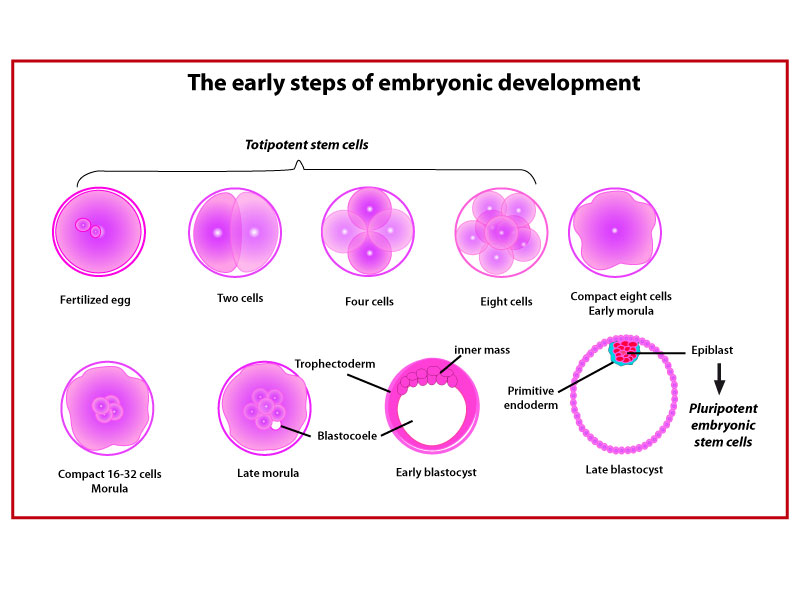
-* The totipotent stem cells are the cells of the embryo from the fertilized egg stage to the eight-cell stage; they can differentiate into all the embryonic and extraembryonic cell types and construct a complete, viable organism.
-* The pluripotent stem cells are the daughter cells of the totipotent cells and can differentiate into all the types of cells with the exception of the extraembryonic cell types; they can give cells derived of any of the three germ layers (endoderm, ectoderm, and mesoderm). Pluripotency is a property of the cells that are located within the inner cell mass of the developing blastocyst. Pluripotency in vivo and in vitro is governed by a trinity of nuclear regulators, Oct4 , Sox2 , and Nanog .
-* The multipotent stem cells can give different types of cell but only those issued from a specific cell lineage.
-* The unipotent stem cell can differentiate into only one type of cell but they can also self-renew as all the stem cells and have to be distinguished from non-stem cells.
Stem cell types are also distinguished according to their tissue origin :
2. Embryonic stem cells
Human embryonic stem cells were first derived from human blastocysts by Thomson et al in 1998. Embryonic stem cells are cells derived from the epiblast of the inner mass of a blastocyst or earlier morula stage embryo. They have the ability to proliferate robustly and are pluripotent cells which have the ability to differentiate in more than 200 different cell types derived of any of the three germ layers (endoderm, ectoderm, and mesoderm).
Embryonic stem cells have been proposed for medical treatments in a wide range of conditions, including degenerative conditions, wound healing, and genetic diseases, in combination with gene therapy.
However, there are major obstacles with the use of human ES cells such as bioethical issues surrounding the destruction of an embryo and immune rejection due to alloantigens following transplantation. As we will see below, other alternatives that do not involve human embryos, such as, adult stem cells, amniotic stem cells, and induced stem cells, are presently proposed to be a substitute to embryonic stem cells.
3. Fetal stem cells
Fetal stem cells are stem cells present in most of the fetal tissues; they are multipotent, generally tissue- specific and they generate the cells of the particular organ or tissue where they are localized.
4. Amniotic fluid-derived stem cells
Amniotic stem cells are multipotent stem cells found in the amniotic fluid. They are considered as an intermediate between the ES cells and the adult stem cells since they express cell markers of both cell lines. They are multipotent and can differentiate into a large number of tissues such as, bone, cartilage, cardiac tissue, nerves, muscle, and skin and also into embryonic cell lines. They have a very high proliferative potential, remaining phenotypically and genetically stable for more than 250 divisions. They do not induce tumors after transplantation. They constitute only1 to 2 % of amniotic fluid cells.(#Antonucci et al, 2011)
5. Adult or somatic stem cells
Adult or somatic stem cells, also named germline stem cells have been identified in almost every organ tested: skin, intestine, bone and bone marrow, liver, heart, brain, pancreas, etc... and can be found in children and adult.
Adult stem cells provide constant replacement cells for tissue homeostasis and repair while, at the same time, maintaining a pool of stem cells by the process of self-renewal, where following cell division, at least one daughter cell is still a stem cell whereas the other is either a stem cell (symmetric division) or a differentiated progeny (asymmetric division).
Most of them are multipotent (lineage-restricted) stem cells which can generate different types of cell only issued from a specific cell lineage. They are referred according to their tissue origin: mesenchymal stem cell, endothelial stem cell, adipose-derived stem cells, dermal stem cells, keratinocyte stem cell, melanocyte stem cell, etc.
Three major classes of adult stem cells are isolated from the bone marrow: hematopoietic stem cells (HSCs), mesenchymal stem cells (MSCs), and multipotent adult progenitor cells (MAPCs).
Haematopoietic stem cells (HSCs) are a heterogeneous group of multipotent stem cells that have the ability to self-renew and differentiate into all the functional blood cell types of the body, including erythrocytes, platelets, and white blood cells. Additionally, HSCs may also give rise to fibrocytes and endothelial progenitor cells. A common feature of hematopoietic lineage cells is the expression of the cell surface antigen CD 45 with the exception of mature red blood cells and their immediate progenitors. The bone marrow is the main site of adult haematopoiesis, although during times of stress, haematopoiesis may also occur in the spleen and liver.(for more details see Wikipedia Hematopoietic stem cell and #Lilly et al., 2011
Mesenchymal stem cells (MSCs), also referred as bone marrow stromal stem cells or marrow stem cells, derived from the adherent stromal cell compartment of adult bone marrow and other tissues, including adipose tissue. It has been shown that they contribute to a significant proportion of cells in the skin although present as a rare cell population in the bone marrow, representing about 0,001% to 0,01% of the nucleated cells, about 10-fold less abundant than hematopoietic stem cells. Recent studies suggest that, in the skin, the bone marrow contributes not only to the inflammatory cells but also to non inflammatory cells, present primarily in the dermis as fibroblast-shaped cells and in the epidermis with a keratinocyte phenotype(). They actively participate to the repair/regeneration of a variety of injured tissues including myocardium, cardiac valves, bone, cartilage, tendon, meniscus, and skin. By definition, mesenchymal stem cells are lineage-negative and express CD105, CD73, and CD90. They differentiate to, at least, osteoblasts, adipo-cytes, and chondroblasts in vitro and are plastic-adherent. (for more details see )
In 2001, the laboratory of Catherine Verfaillie at the University of Minnesota described the multipotent adult progenitor cell (MAPC) as a novel progenitor non hematopoietic stem cell present in adult bone marrow that can be co-purified with mesenchymal stem cell from BM cells but is biologically and antigenically distinct from the mesenchymal stem cell (#Reyes and Verfaillie, 2001). MAPCs express the pluripotent state-specific transcription factors Oct-3/4 and Rex-1, and can differentiate into cell types representative of all 3 germ layers; thus, MAPCs are generally believed to be a more primitive cell type than MSCs. MAPCs thus embody a unique class of adult stem cells that exhibit the broad biological plasticity characteristic of embryonic stem (ES) cells, while maintaining the characteristics that make adult stem cells more amenable to therapeutic appli-cation. MSCs kept for prolonged periods in culture tend to lose their differentiation capabilities and undergo senescence at approximately 20 to 40 population doublings. In contrast to MSCs, MAPCs have an average telomere length that remained constant for up to 100 population doublings in vitro. Based upon their differential potential and reduced senescence, MAPCs have been considered as a potentially desirable non hematopoietic stem cell source for use in allogeneic bone marrow transplantation.
6. Induced pluripotent stem cells or iPS cell
Induced pluripotent stem cells or iPS cell have been discovered in 2006, by Takahashi and Yamanaka who showed how to “reprogram” mouse cells with a specialized function (for example, skin cells) in the laboratory, so that they behave like an embryonic stem cell. These iPS cells were created by inducing the specialized cells to express genes that are normally made in embryonic stem cells and that control how the cell functions.
For this purpose, the scientists used the retrovirus-mediated transduction of 4 transcription factors (c-Myc, Oct3/4, SOX2, and Klf4 ) into mouse fibroblasts to generate iPS cells that were selected by the expression of a -geo cassette (a fusion of the galactosidase and neomycin resistance genes) driven by the mouse Fbx15 promoter. These Fbx15 iPS cells, when subcutaneously injected into nude mice, gave rise to teratomas with histological evidence of cells differentiating into all 3 germ layers. However, Fbx15 iPS cells showed different gene expression and DNA methylation patterns compared with ES cells and failed to contribute to adult chimeras.
In 2007, human iPS cells were established by the transduction of either the same set of transcriptional factors (c-Myc, Oct3/4, SOX2, Klf4) or another set of transcriptional factors (Oct3/4, SOX2, Nanog, Lin28) into human fibroblasts. These human iPS cells are similar to human ES cells in their morphology, their gene expression, and the epigenetic status of pluripotent cell specific genes, and telomerase activity. Furthermore, these cells can differentiate into the cell types of the 3 germ layers in vitro and form teratomas in vivo.
To obtain high quality cells, it was later shown that the expression of either Nanog or Oct3/4, which are closely involved in pluripotency, has to be used to select the cells; such cells contribute to adult chimeras and show germline transmission. Unfortunately, c-Myc, one of the four genes used for the reprogrammation is an oncogene and 20 % of the chimeric mice developed cancers. Re-programmation without c-Myc was possible but was not as efficient. It has been recently discovered by #Maekawa et al. that Gli-like transcription factor Glis1 associated with the transcription factors Oct3/4, Sox2, and Klf4 remarkably enhanced iPSC generation from mouse and human fibroblasts without increasing the transformation of these cells.
The potential of iPS cells is having a significant impact on regenerative medicine, with the promise of infinite self-renewal, differentiation into multiple cell types, and no problems concerning ethics or immunological rejection. The production of iPS gives a novel cell source of pluripotent stem cells circumventing the ethical issues associated with generating ES cells; the sacrifice of an embryo is dispensable, and the use of the recipient’s own cells eliminates the risk of immune rejection generally associated with transplantation.
Generally, it is difficult or impossible to take large samples from specific patient tissues or such terminally differentiated cells cannot proliferate sufficiently in vitro and provide enough tissue for being further studied. Therefore, another expectation for iPS cells is the generation of human disease-specific pluripotent stem cells, since established iPS cells have identical genetic mutations to the source tissue. It has been shown that patient-specific iPS cells can differentiate into any type of cell, allowing to directly and repetitively analyze the diseased cells in vitro and furthermore to study the effects of known drugs and to screen new compounds in combination with personalized medicine.
To date, patient-specific iPS cells have been generated to diseases such as adenosine deaminase deficiency, the Schwachman-Bodian-Diamond syndrome, Gaucher’s disease, the Duchenne and Becker muscular dystrophy, Parkinson’s disease, Huntington’s disease, type 1 diabetes mellitus, Down’s syndrome, the Lesch-Nyhan syndrome, amyotrophic lateral sclerosis, spinal muscular atrophy, and Fanconi’s anemia.
7. iPS cells and skin
Concerning skin, in addition to fibroblasts, it has been demonstrated that epidermal keratinocytes and hair follicle outer root sheath cells from adults can be reprogrammed to iPS cells.
More recently, three papers have reported that mouse and human iPS cells can be differentiated into keratinocytes through the sequential application of retinoic acid to promote ectodermal fate and bone morphogenetic protein 4 (BMP4) to block neural fate(#Bilousova et al., 2011; #Tolar et al., 2011; #Itoh et al., 2011). The stem cell population could be enriched by the selective attachment to type IV collagen-coated surfaces, onto which the fully differentiated progeny does not attach.
Itoh et al. succeeded:
-* in generating iPS cells from normal human fibroblasts but also from fibroblasts isolated from patients with recessive dystrophic epidermolysis bullosa ,
-* in differentiating both of these iPS cells into keratinocytes
-* and in generating 3D skin equivalents.
The iPSC-derived keratinocytes expressed not only K14, but also other keratinocyte markers, including p63 (mainly ΔNp63), DSG3, ITGB4, laminin 5, and keratin 5. Additionally, keratin 18, a marker of other epithelial cells that are not expressed in the skin, was similarly absent in these cells. As expected, type VII collagen was expressed in normal iPSC-derived K14+ cells, but not in the RDEB patient-specific iPSC-derived K14+ cells. Moreover, after culturing the induced cells in high calcium media to induce in vitro epidermal differentiation, they expressed keratin 1 (K1), a marker of epidermal differentiation, indicating that these iPSC-derived keratinocytes closely resemble normal human keratinocytes.
8. Induced differentiated cells
Recently, a new alternative pathway has been proposed to produce differentiated cells from a large numbers of patients. This consists in a process named transdifferentiation which is a direct reprogramming of differentiated cells in another type of differentiated cells, called induced differentiated cells, without transient dedifferentiation to a progenitor or embryonic cell state.
Thus, it was shown that somatic cells can also be directly converted to other mature cell types by expression of a specific combination of genes. In 2008, #Zhou et al showed that pancreatic exocrine cells could be converted in vivo to pancreatic β-cells by infecting them with adenovirus expressing three transcription factors, Ngn3(=Neurog3), Pdx1 and Mafa, all known to be important for β-cell development. More recently, in 2010, #Vierbuchen et al., 2010 showed that mouse fibroblasts, following treatment with lentivirus containing genes for three transcription factors expressed in the nervous system, Ascl1, Brn2, and Myt1l, could differentiate directly into neurons. Later, over-expression of the same three transcription factors was shown to also convert, human embryonic and postnatal fibroblasts to functional neurons (#Pfisterer et al., 2011. In addition, expression of a minimal set of three transcription factors, Mash1 (also known as Ascl1), Nurr1 (also known as Nr4a2) and Lmx1a is able to generate directly functional dopaminergic neurons(Induced dopaminergic (iDA) cells) from mouse and human fibroblasts without reverting to a progenitor cell stage (#Caiazzo et al., 2011). Also in 2010, it has been reported that post-natal cardiac or dermal fibroblasts may be directly reprogrammed into cardiomyocyte-like cells by a combination of three developmental transcription factors (Gata4, Mef2c and Tbx5) (#Ieda et al. 2010). Moreover, #Szabo et al., 2010 showed that the ectopic expression of OCT4-activated haematopoietic transcription factors, together with specific cytokine treatment, could induce the transdifferentiation of fibroblasts into blood cells.
In conclusion, the cells generated via direct conversion do not pass through a pluripotent state, are probably not tumorigenic, and may therefore serve as an interesting alternative to iPS cells for generating patient- and/or disease-specific cells.
Bibliography
Reviews
Bianco P, Robey PG, Simmons PJ. Mesenchymal stem cells: revisiting his-tory, concepts, and assays. Cell Stem Cell. 2008 Apr 10;2(4):313-9. PubMed PMID: 18397751; PubMed Central PMCID: PMC2613570.
Lilly AJ, Johnson WE, Bunce CM. The haematopoietic stem cell niche: new insights into the mechanisms regulating haematopoietic stem cell behaviour. Stem Cells Int. 2011;2011:274564. Epub 2011 Oct 30. PubMed PMID: 22135682; PubMed Central PMCID: PMC3205662.
Wu Y, Zhao RC, Tredget EE. Concise review: bone marrow-derived stem/progenitor cells in cutaneous repair and regeneration. Stem Cells. 2010 May;28(5):905-15. Review. PubMed PMID: 20474078; PubMed Central PMCID: PMC2964514.
For further details on iPS cell production, see:
References
Antonucci I, Stuppia L, Kaneko Y, Yu S, Tajiri N, Bae EC, Chheda SH, Weinbren NL, Borlongan CV. Amniotic fluid as a rich source of mesenchymal stromal cells for transplantation therapy. Cell Transplant. 2011;20(6):789-95. Epub 2010 Nov 5. PubMed PMID: 21054947.
Bilousova G, Chen J, Roop DR. Differentiation of mouse induced pluripotent stem cells into a multipotent keratinocyte lineage. J Invest Dermatol. 2011 Apr;131(4):857-64. Epub 2010 Dec 9. PubMed PMID: 21150926.
[Caiazzo et al., 2011] Caiazzo M, Dell’Anno MT, Dvoretskova E, Lazarevic D, Taverna S, Leo D, Sotnikova TD, Menegon A, Roncaglia P, Colciago G, Russo G, Carninci P, Pezzoli G, Gainetdinov RR, Gustincich S, Dityatev A, Broccoli V. Direct generation of functional dopaminergic neurons from mouse and human fibroblasts. Nature. 2011 Jul 3;476(7359):224-7. doi: 10.1038/nature10284. PubMed PMID: 21725324.
Itoh M, Kiuru M, Cairo MS, Christiano AM. Generation of keratinocytes from normal and recessive dystrophic epidermolysis bullosa-induced pluripotent stem cells. Proc Natl Acad Sci U S A. 2011 May 24;108(21):8797-802. Epub 2011 May 9. PubMed PMID: 21555586; PubMed Central PMCID: PMC3102348.
Ieda M, Fu JD, Delgado-Olguin P, Vedantham V, Hayashi Y, Bruneau BG, Srivastava D. Direct reprogramming of fibroblasts into functional cardiomyocytes by defined factors. Cell. 2010 Aug 6;142(3):375-86. PubMed PMID: 20691899; PubMed Central PMCID: PMC2919844.
Maekawa M, Yamaguchi K, Nakamura T, Shibukawa R, Kodanaka I, Ichisaka T, Kawamura Y, Mochizuki H, Goshima N, Yamanaka S. Direct reprogramming of somatic cells is promoted by maternal transcription factor Glis1. Nature. 2011 Jun 8;474(7350):225-9. doi: 10.1038/nature10106. PubMed PMID: 21654807.
[Maherali et al. ,2007]<-]Maherali N, Sridharan R, Xie W, Utikal J, Eminli S, Arnold K, Stadtfeld M, Yachechko R, Tchieu J, Jaenisch R, Plath K, Hochedlinger K. Directly reprogrammed fibroblasts show global epigenetic remodeling and widespread tissue contribution. Cell Stem Cell. 2007 Jun 7;1(1):55-70. PubMed PMID: 18371336.
Okita, K; Ichisaka, T; Yamanaka, S(2007). “Generation of germline-competent induced pluripotent stem cells”. Nature 448 (7151): 313–7.
Pfisterer U, Kirkeby A, Torper O, Wood J, Nelander J, Dufour A, Björklund A, Lindvall O, Jakobsson J, Parmar M. Direct conversion of human fibroblasts to dopaminergic neurons. Proc Natl Acad Sci U S A. 2011 Jun 21;108(25):10343-8. Epub 2011 Jun 6. PubMed PMID: 21646515; PubMed Central PMCID: PMC3121829.
Reyes M, Verfaillie CM. Characterization of multipotent adult pro-genitor cells, a subpopulation of mesenchymal stem cells. Ann N Y Acad Sci. 2001 Jun;938:231-3; discussion 233-5. PubMed PMID: 11458512.
Szabo E, Rampalli S, Risueño RM, Schnerch A, Mitchell R, Fiebig-Comyn A, Levadoux-Martin M, Bhatia M. Direct conversion of human fibroblasts to multilineage blood progenitors. Nature. 2010 Nov 25;468(7323):521-6. Epub 2010 Nov 7. PubMed PMID: 21057492.
Tolar J, Xia L, Riddle MJ, Lees CJ, Eide CR, McElmurry RT, Titeux M, Osborn MJ, Lund TC, Hovnanian A, Wagner JE, Blazar BR. Induced pluripotent stem cells from individuals with recessive dystrophic epidermolysis bullosa. J Invest Dermatol. 2011 Apr;131(4):848-56. Epub 2010 Dec 2. PubMed PMID: 21124339.
Vierbuchen T, Ostermeier A, Pang ZP, Kokubu Y, Südhof TC, Wernig M. Direct conversion of fibroblasts to functional neurons by defined factors. Nature. 2010 Feb 25;463(7284):1035-41. Epub 2010 Jan 27. PubMed PMID: 20107439; PubMed Central PMCID: PMC2829121.
Wernig, M; Meissner, A; Foreman, R; Brambrink, T; Ku, M; Hochedlinger, K; Bernstein, BE; Jaenisch, R (2007). “In vitro reprogramming of fibroblasts into a pluripotent ES-cell-like state”. Nature 448 (7151): 318–24.
Zhou Q, Brown J, Kanarek A, Rajagopal J, Melton DA. In vivo reprogramming of adult pancreatic exocrine cells to beta-cells. Nature. 2008 Oct 2;455(7213):627-32. Epub 2008 Aug 27. PubMed PMID: 18754011.
Site powered by SPIP 3.0.17 + AHUNTSIC

Visitors logged in: 91