L’EPIDERME, UNE BARRIERE SUR TOUS LES FRONTS
La peau constitue l’interface entre l’organisme et l’environnement extérieur. Sa raison d’être est d’assurer une mission de protection en formant une véritable barrière qui est vitale. En effet, qu’elles soient d’ordre physique (facteurs mécaniques, thermiques, rayonnements UV , etc.), chimique (tensioactifs, solvants, allergènes, etc.) ou biologique (agents infectieux), le tissu cutané est quotidiennement soumis à de multiples agressions. En plus de cette fonction d’obstacle à la pénétration d’éléments extérieurs, la peau permet de lutter contre la déshydratation en limitant la diffusion de l’eau hors de l’organisme.
Cette barrière cutanée est en grande partie assurée par la couche la plus superficielle de la peau, l’épiderme . On parle alors de « barrière épidermique ». La fonctionnalité de cette dernière est dépendante de la mise en place de différents systèmes très sophistiqués lui assurant de multiples rôles. Ainsi, l’épiderme joue à la fois un rôle de barrière hydrique, physique, anti-oxydante, photo-protectrice mais également anti-microbienne (#Madison, 2003).

I. Une barrière hydrique
La fonction de barrière hydrique de l’épiderme correspond au maintien d’un gradient de la teneur en eau entre les couches profondes de l’épiderme et la couche cornée . Dans les conditions physiologiques, la surface cutanée contient environ 20% d’eau, alors que l’épiderme profond présente une teneur en eau de l’ordre de 70%. L’homéostasie hydrique est un élément indispensable à l’équilibre physiologique de la peau. En effet, outre son influence sur les paramètres macroscopiques apparents, tels que l’élasticité cutanée, le taux d’hydratation régule également les activités enzymatiques et la signalisation cellulaire au sein de l’épiderme. Le maintien d’un gradient hydrique optimal est assuré par les lipides intercornéocytaires , les jonctions serrées ainsi que par les composants du Facteur Naturel d’Hydratation (FNH) (NMF en anglais pour « Natural Moisturizing Factor »)
.
I.1. Les lipides intercornéocytaires
Les cornéocytes sont entourés de lipides organisés en feuillet qui jouent un rôle fondamental de barrière hydrophobe dynamique (#van Smeden et al, 2014 ; #Feingold and Elias, 2013). Ils sont le siège de séquences d’absorption et de désorption des molécules d’eau au sein des espaces intercornéocytaires. Essentiellement synthétisés par les kératinocytes et issus de la sécrétion des corps lamellaires (voir L’épiderme et la différenciation des kératinocytes) , les lipides de la couche cornée présentent une composition bien spécifique. Les principaux lipides présents dans la couche cornée sont les céramides (45-50 % en masse), le cholestérol (25 % en masse) et les acides gras libres (10-15 % en masse). Ces différentes espèces sont présentes en quantités approximativement équimolaires. On retrouve également 5-10 % d’autres espèces lipidiques comme des phospholipides et du sulfate de cholestérol impliqué dans la régulation de la desquamation. La composition et l’organisation inhabituelles de ces lipides semblent spécifiquement adaptées pour limiter la pénétration des molécules non liposolubles à travers la couche cornée. Il est important de noter que la composition lipidique de la couche cornée varie selon l’âge, le sexe, le territoire anatomique et les saisons. Les espaces intercornéocytaires les plus superficiels contiennent également des triglycérides et des cires qui recouvrent la peau et sont sécrétés par les glandes sébacées.
I.1.1. Les différentes espèces de lipides intercornéocytaires
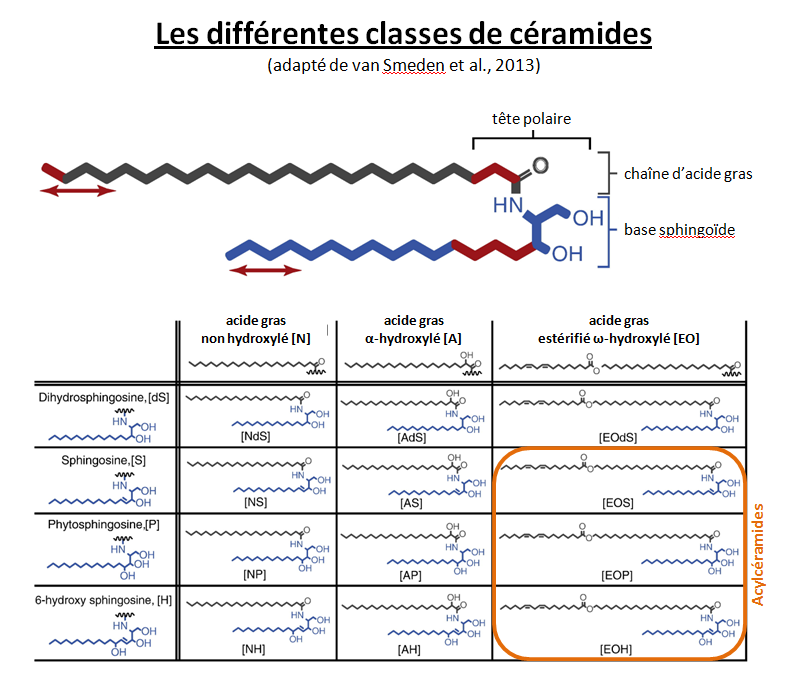
Les céramides constituent une famille de lipides structurellement hétérogènes et complexes. Leur squelette est constitué d’une base sphingoïde liée à un acide gras par une liaison amide. Selon leur structure chimique, les bases sphingoïdes peuvent être de quatre types : sphingosine (S), dihydrosphingosine (DS), phytosphingosine (P) et 6-hydroxy sphingosine (H). L’acide gras peut quant à lui être non hydroxylé (N), α-hydroxylé (A) ou estérifié ω-hydroxylé (EO) avec de l’acide linoléique. À ce jour, les CER[NDS], CER[NS], CER[NP], CER[NH], CER[ADS], CER[AS], CER[AP], CER[AH], CER[EOS], CER[EOP] et CER[EOH] ont été identifiés comme faisant partie intégrante des lipides intercornéocytaires. Parmi ces différentes classes de CER de la couche cornée, les CER[EOS], CER[EOP] et CER[EOH], constituant les acylcéramides, présentent une structure moléculaire remarquable avec une longueur de chaîne de 28 à 32 carbones alors que la longueur moyenne des autres céramides est de 26 à 28 carbones (à l’exception du céramide AS qui a une chaîne moyenne de C15-C18). En effet, un acide linoléique est lié à un ω-hydroxy-acide gras à longue chaîne par une fonction ester. Cette longueur de chaîne importante permet aux acylcéramides de s’ancrer à l’enveloppe cornée au cours de sa formation grâce à la transglutaminase 1 catalysant la formation d’une liaison covalente avec l’involucrine . Ces acylcéramides, formant une sorte de « treillis » de molécules imbriquées les unes dans les autres, servent d’échafaudage pour la mise en place des lamellae lipidiques. La synthèse des céramides résulte de 2 voies principales et est fonction de l’espèce considérée. La première voie repose sur le catabolisme des précurseurs lipidiques véhiculés par les corps lamellaires. Les glucosylcéramides et les sphingomyélines sont dégradés par, respectivement, la β-glucocérébrosidase et la sphingomyélinase. La deuxième voie correspond à une synthèse de novo à partir de la condensation de sérine et de palmitoyl-CoA par deux enzymes, la sérine palmitoyltransférase et la céramide synthase.
Le cholestérol est néosynthétisé dans les couches profondes de l’épiderme à partir d’acétate par la voie de l’hydroxyméthyl-coenzyme A réductase. Toutefois, une petite proportion est issue de la réabsorption par les kératinocytes basaux à partir de la circulation sanguine. Le cholestérol peut ensuite être converti en sulfate de cholestérol par la sulfotransférase au sein de la couche granuleuse. Alors que le transfert du cholestérol vers la couche cornée passe par la sécrétion des corps lamellaires, le caractère hydrophobe du sulfate de cholestérol lui permet de franchir la membrane plasmique pour rejoindre l’espace intercornéocytaire. Au sein de la couche cornée, le sulfate de cholestérol est à nouveau converti en cholestérol sous l’action de la stéroïde sulfatase.
La couche cornée contient une trentaine d’acides gras libres différents. Majoritairement saturés, ils présentent de longues chaînes carbonées (C14 à C28). Les acides gras libres principaux sont les acides palmitique (C16:0), stéarique (C18:0), oléique (C18:1ω9), béhénique (C22:0) et lignocérique (C20:0). Cette composition unique en acide gras libres provient presque exclusivement d’une production de novo dans les espaces intercornéocytaires via l’hydrolyse des glycérophospholipides par des phospholipases A2 (PLA2) sécrétées.
I.1.2. L’organisation des lipides intercornéocytaires
Les lipides intercornéocytaires présentent un arrangement en lamellae superposées. Ils sont ainsi disposés perpendiculairement à la surface des cornéocytes, parallèles les uns aux autres, de part et d’autre des enveloppes cornées adjacentes.

En plus de cette organisation lamellaire, les lipides de la couche cornée présentent un assemblage latéral bien organisé (#Bouwstra et al, 2001). La majorité des lipides de la couche cornée sont organisés de manière très compacte. On parle d’état cristallin ou d’arrangement orthorhombique. Cet empilement resserré des lipides renforce l’imperméabilité de la couche cornée. En remontant vers la surface de la couche cornée, se produit une fluidification des lipides qui se retrouvent plutôt dans un état de gel (arrangement hexagonal) voire même liquide.
L’intégrité de la structure des lipides intercornéocytaires est en partie assurée par les céramides. En effet, ces lipides, qui possèdent un haut point de fusion, se présentent, à température ambiante, dans un état cristallin. De plus, les acylcéramides, notamment les CER[EOS], favoriseraient la compacité des lamellae lipidiques. Quant au cholestérol qui interagit pour partie avec les longues chaînes d’acides gras des céramides, il assurerait une certaine flexibilité de la couche cornée. Les acides gras libres s’intercalent entre les phases lamellaires et sont essentiels à la mise en place de la structure latérale des lipides.
Bien que les trois espèces de lipides de la couche cornée présentent un rôle bien défini, la barrière hydrique résulte surtout d’un équilibre compositionnel et d’une synergie entre ces différents lipides. L’organisation en lamellae, la compaction et la nature hydrophobe des lipides intercornéocytaires permettent d’assurer l’étanchéité de la couche cornée pour limiter le flux de solutions aqueuses depuis l’intérieur de l’organisme vers l’extérieur et inversement. Toutefois, cette imperméabilité n’est pas totale. On note la présence d’une perte insensible en eau (en moyenne 9 g par m2 de peau par heure) dans le sens « intérieur vers extérieur » permettant le maintien d’un gradient hydrique physiologique.
I.2. Les jonctions serrées
La barrière hydrique est également assurée par des jonctions intercellulaires spécifiques des épithéliums appelées jonctions serrées (#Kirschner and Brandner, 2012). Dans l’épiderme, ces dernières sont localisées au niveau de la membrane apico-latérale des kératinocytes granuleux. Observées en microscopie électronique après cryofracture, elles forment une ceinture assurant un contact extrêmement étroit entre deux cellules adjacentes. Outre leur rôle organisationnel dans la polarisation cellulaire en bloquant la diffusion libre des lipides et protéines membranaires entre le pôle basolatéral et le pôle apical, les jonctions serrées assurent une étanchéité presque totale de l’épiderme. En effet, elles permettent de limiter le passage des solutés par l’espace intercellulaire (barrière paracellulaire) en ne laissant passer que les petites molécules et les ions et assurent donc un rôle de pore sélectif intercellulaire.
Ces structures jonctionnelles présentent une structure complexe. Elles sont constituées de protéines transmembranaires : l’occludine, les claudines (plus de 24 isoformes identifiés chez l’Homme), les « Junctional Adhesion Molecule » (JAM) et la tricelluline. Ces protéines permettent la fusion des feuilles externes des membranes de 2 cellules adjacentes de par la mise en place d’interactions homophiles. Ces protéines transmembranaires sont reliées au cytosquelette d’actine via des protéines d’échafaudage cytoplasmique sous-membranaire. Il s’agit des protéines ZO-1, ZO-2 et ZO-3 (pour zonula occludens proteins) à domaines PDZ, des protéines MUPP1 (Multi-PDZ Protein 1) et de la cinguline.
Le rôle fonctionnel des jonctions serrées dans l’étanchéité de l’épiderme a été démontré grâce à un modèle murin déficient pour la claudine 1 (#Furuse et al, 2002). En effet, l’absence de cette protéine est à l’origine d’une létalité précoce due à une déshydratation massive. En plus d’une couche cornée extrêmement compacte, ces souris présentent des jonctions serrées non fonctionnelles à l’origine de la déshydratation.
Outre ce rôle majeur dans l’imperméabilité de l’épiderme, les jonctions serrées peuvent également constituer un système de compensation lors de l’altération de la barrière épidermique. En effet, en cas d’ichtyose vulgaire, de psoriasis ou au cours de la cicatrisation cutanée, on retrouve une expression de certaines protéines des jonctions serrées telles que ZO1 et l’occludine, dans les couches profondes de l’épiderme alors qu’elle est d’ordinaire restreinte à la couche granuleuse.
I.3. Le Facteur Naturel d’Hydratation
Le Facteur Naturel d’Hydratation (FNH) est spécifique de la couche cornée où on le retrouve en grande quantité puisqu’il en représente jusqu’à 30% du poids sec total. Il est composé, à plus de 50%, d’acides aminés libres et de leurs dérivés tels que l’acide carboxylique pyrrolidone dérivé de la glutamine et l’acide urocanique dérivé de l’histidine (#Rawlings and Matts, 2005). A ces acides aminés viennent s’ajouter des composants extracellulaires tels que l’acide lactique, l’urée et les sucres. Alors que ces derniers composants sont issus de la sécrétion séborrhéique par les glandes sébacées, les acides aminés sont majoritairement issus de la protéolyse d’une protéine unique, la filaggrine .
La filaggrine présente un métabolisme complexe.

Elle est synthétisée sous la forme d’un précurseur de grande taille, la profilaggrine, qui va être phosphorylé et stocké dans le cytoplasme des kératinocytes granuleux sous la forme de granules de kératohyaline. Ce précurseur de haut poids moléculaire est composé de 10 à 12 sous-unités de filaggrine répétées en tandem. Lors du processus de cornification, la profilaggrine est déphosphorylée puis protéolysée en sous-unités monomériques de filaggrine. Celles-ci s’associent aux filaments intermédiaires de kératines permettant la formation de la matrice fibreuse intra-cornéocytaire. Au sein des assises cellulaires profondes de la couche cornée, ces sous-unités subissent une modification post-traductionnelle appelée désimination (ou encore citrullination) : les résidus arginyl sont transformés en résidus citrullyl par une (ou des) peptidylarginine désiminase(s), engendrant un changement global de charge. La filaggrine désiminée, alors acide, perd son affinité pour les filaments intermédiaires de kératines avant d’être complètement dégradée, générant les acides aminés qui constituent le FNH.
Le FNH présente des propriétés hygroscopiques. Il est capable de capter et de retenir l’eau au sein de la matrice intracornéocytaire. Il permet ainsi d’assurer l’hydratation de la couche cornée, propriété essentielle à sa flexibilité et à son activité métabolique.
II. Une barrière physique
La résistance mécanique de l’épiderme est conférée par la mise en place d’une enveloppe cornée rigide renfermant une matrice intracornéocytaire dense. Les enveloppes cornées adjacentes sont étroitement liées entre elles par des systèmes jonctionnels spécifiques, les cornéodesmosomes. C’est donc la continuité entre ces trois composants, cornéodesmosomes, enveloppe cornée et matrice fibreuse qui permet la mise en place d’une barrière épidermique physique.
II.1. L’enveloppe cornée
Lors des étapes terminales de la différenciation épidermique, les kératinocytes subissent d’importants changements morphologiques avec notamment le remplacement de la membrane plasmique par une coque protéique rigide et insoluble, l’enveloppe cornée. Au niveau interne, la structure protéique est liée aux composants de la matrice fibreuse par des liaisons covalentes. Au niveau externe, elle est recouverte d’une structure lipidique constituée d’une monocouche d’acylcéramides dont l’une des extrémités interagit avec les lipides des espaces intercornéocytaires.

L’enveloppe cornée se met en place progressivement (#Steinert and Marekov, 1995 ; #Candi et al, 2005). La phase d’initiation a lieu sous la membrane plasmique des kératinocytes épineux avec l’assemblage covalent de plusieurs précurseurs protéiques grâce à l’action de différentes transglutaminases. Ces enzymes catalysent la formation de liaisons covalentes entre les résidus glutamine et lysine des protéines. Il y a tout d’abord mise en place d’un réseau covalent de périplakine, d’envoplakine et d’involucrine formant une véritable armature protéique, qui s’étend progressivement depuis la plaque desmosomale sur toute la face interne de la membrane plasmique. Dans un deuxième temps, se met en place une monocouche lipidique au niveau de la face externe. Les acylcéramides, issus de la sécrétion des corps lamellaires, sont liés de façon covalente par des liens esters à l’échafaudage protéique déjà en place par l’action de la transglutaminase 1. Ils remplacent ainsi progressivement les phospholipides de la membrane plasmique. De façon concomitante, la structure protéique primaire est renforcée par la liaison covalente d’autres protéines telles que la loricrine , les protéines « Small PRolin Rich », des protéines de la famille S100, les protéines « late-cornified envelope », l’hornerine, etc. Finalement, cette coque protéique est liée de manière covalente aux filaments de kératines et à la filaggrine. Au cours de sa mise en place, l’enveloppe cornée englobe une partie des composants intracellulaires et transmembranaires des desmosomes .
L’enveloppe cornée permet, de par son échafaudage protéique, la séquestration des macrofibrilles de kératines de la matrice cornéocytaire et, de par sa structure lipidique, l’ancrage aux lipides intercornéocytaires. Ainsi, cette structure hautement résistante contribue aux propriétés biomécaniques de la couche cornée.
II.2. La matrice fibreuse intracornéocytaire
La résistance aux contraintes mécaniques que subit l’épiderme est, en partie, assurée par le cytosquelette des cornéocytes constitué d’un réseau de filaments intermédiaires composé de kératines. Ces dernières constituent les protéines structurales majeures de l’épiderme et représentent environ 85% des protéines totales de la couche cornée. D’un point de vue moléculaire, on distingue les kératines de type 1 (kératines acides) et les kératines de type 2 (kératines basiques). Elles sont capables de former des hétérodimères qui, en se polymérisant, vont donner naissance aux filaments intermédiaires. Dans la couche cornée, les filaments de kératines s’agrègent grâce à la formation de liaisons ioniques, sous l’action de la filaggrine (pour filament aggregating protein), membre de la famille des protéines associées aux filaments intermédiaires (#Brown and McLean, 2013). Les kératines sont aussi liées entre elles par des ponts disulfures. Cette agrégation des kératines en macrofibrilles permet de former la matrice fibreuse intracornéocytaire extrêmement dense et indispensable à la résistance mécanique de la couche cornée.
II.3. Les desmosomes et cornéodesmosomes

La fonction de barrière physique assurée par l’épiderme repose également sur la présence de jonctions intercellulaires appelées desmosomes. Il s’agit de structures d’ancrage pour les filaments intermédiaires du cytosquelette. Ils assurent la cohésion entre deux cellules adjacentes à la manière d’un système de « bouton pression » et forme ainsi un réseau trans-cellulaire qui confère aux kératinocytes leur résistance et leur extensibilité. Les observations en microscopie électronique ont permis de révéler leur structure symétrique complexe. Au centre, dans l’espace intercellulaire, on trouve le cœur protéique qui est entouré, au niveau de la face interne de la membrane plasmique, par deux plaques discoïdes cytoplasmiques, auxquelles viennent s’ancrer les filaments intermédiaires de kératines.
Le cœur desmosomal est composé de la partie extracellulaire de cadhérines, glycoprotéines transmembranaires dépendantes du calcium divisées en 2 familles : les desmocollines et les desmogléines. Il existe quatre gènes différents codant pour les desmogléines et trois codant pour les desmocollines, tous regroupés dans un même cluster sur le chromosome 18. De par leur domaine intracellulaire carboxy-terminal, les cadhérines desmosomales interagissent avec les protéines de la plaque desmosomale. Ces dernières appartiennent à la superfamille des plakines (desmoplakines, périplakine et envoplakine) et à celle des protéines à domaine armadillo (plakoglobine et plakophilines). Les protéines de la plaque desmosomale s’associent aux filaments intermédiaires de kératines et permettent la liaison indirecte de ces derniers aux protéines du cœur desmosomal.
L’expression des différents isoformes de protéines desmosomales varie au cours de la différenciation épidermique et ce en même temps que le nombre et la taille des desmosomes augmentent. La formation de ces structures, dépendante de l’interaction entre les cadhérines et les filaments de kératines, est régulée par le calcium ainsi que par différentes kinases et phosphatases membranaires.
Au cours de la cornification, les desmosomes subissent des modifications structurales majeures pour former les cornéodesmosomes. La plaque desmosomale est progressivement incorporée à l’enveloppe cornée jusqu’à devenir invisible en microscopie électronique et le cœur intercellulaire se densifie. Ceci s’accompagne de l’intégration d’une protéine supplémentaire, la cornéodesmosine . Cette protéine est synthétisée au sein des kératinocytes épineux supérieurs. Sécrétée par l’intermédiaire des corps lamellaires, la cornéodesmosine est secondairement incorporée au niveau du cœur des desmosomes. Il s’agit de la seule protéine identifiée à ce jour comme étant spécifique des cornéodesmosomes. Son contenu riche en résidus sérine et glycine s’organisant en boucles souples est compatible avec des fonctions d’adhésion. Toutefois, ses partenaires protéiques restent encore à définir.
III. Une barrière anti-oxydante
En contact direct avec l’environnement extérieur, la peau constitue une cible privilégiée du stress oxydatif (#Vermeij et al, 2011). En effet, il existe de nombreux facteurs exogènes oxydants parmi lesquels les agents polluants, les radiations ionisantes et non-ionisantes. La peau subit également l’effet pro-oxydant de facteurs endogènes tels que les réactions chimiques de divers systèmes enzymatiques ou secondaires à l’inflammation (psoriasis ou autres pathologies inflammatoires cutanées). L’action de ces différents facteurs se traduit par la production d’espèces réactives de l’oxygène (ROS), également dénommées dérivés réactifs de l’oxygène, qui se forment secondairement à une réduction incomplète de l’oxygène. Ces composés participent à des réactions d’oxydation physiologiquement indispensables mais peuvent, dans certaines conditions, être également nuisibles. En effet, ils peuvent être responsables de réactions d’oxydation irréversibles à l’origine de cassures et mutations de l’ADN, d’inactivations des protéines et de peroxydations lipidiques. Au niveau cutané, cela se traduit sous différentes formes : altération de l’élasticité tissulaire, vieillissement précoce, inflammation, anomalies de kératinisation ou encore développement de cancer .
La mise en place de multiples systèmes de défense contre l’activité des ROS au sein de l’épiderme est donc cruciale. Il peut s’agir de mécanismes de prévention bloquant leur formation ainsi que de mécanismes de réparation ou d’élimination des cibles biologiques altérées. Ces systèmes sont de nature enzymatique (superoxyde dismutase , catalase , peroxydase du glutathion,…) ou moléculaire. Les antioxydants moléculaires correspondent à des réducteurs endogènes (le gluthathion, l’acide urique , la mélanine ,…) ou exogènes (l’acide ascorbique , l’α-tocophérol, les polyphénols,…). Certains des composants de l’enveloppe cornée en particulier les protéines SPRR, sont aussi capables de désactiver les ROS. voir Le stress oxydant cutané
IV. Une barrière photo-protectrice
La peau constitue la première cible des rayonnements UV . Ces derniers peuvent induire divers dommages cutanés qui, à long terme, sont à l’origine du photo‐vieillissement et de la photo-carcinogénèse cutanée (voir Effets des UV sur l’ADN : lésions et mutations) (voir Effets tissulaires des UV). Plusieurs mécanismes de photo-protection sont mis en place au sein de l’épiderme où les rayonnements UV peuvent être réfléchis ou absorbés. L’absorption des UV induit, en effet, l’activation de nombreuses voies de signalisation cellulaire et la mise en place de plusieurs réponses au niveau tissulaire.
IV.1. La structure de la couche cornée
Une exposition aux UV induit une augmentation du nombre d’assises de kératinocytes (acanthose) ainsi que de cornéocytes (hyperkératose). Bien que son rôle photoprotecteur reste modeste, cet épaississement de l’épiderme permet de limiter la pénétration des rayonnements. De plus, cet épaississement de l’épiderme permet de limiter la pénétration des rayonnements en favorisant leur absorption par les kératines.
IV.2. L’acide urocanique
L’action protectrice des pigments de mélanine est complétée par celle de l’acide urocanique, un dérivé de l’histidine libre retrouvé en forte concentration dans la couche cornée. Cette histidine libre est issue de la protéolyse de la filaggrine et subit une désamination non-oxydative catalysée par l’histidine ammonia-lyase (ou histidase) permettant la formation de l’acide trans-urocanique. Celui-ci constitue un véritable filtre solaire endogène. En effet, il est capable d’absorber les UV-B en subissant une photo-isomérisation en acide cis-urocanique. Toutefois, l’acide urocanique joue également un rôle dans la photo-carcinogenèse. Il participe en effet à l’immunosuppression induite par les UV en activant les lymphocytes T régulateurs et en inhibant les cellules de Langerhans .
IV.3. La mélanogenèse
La mélanogenèse, processus de synthèse des mélanines, pigments naturels de la peau et des poils, constitue le système photo-protecteur majeur. Les mélanines, eumélanines (pigment sombres) et phaeomélanines (pigments claires), sont issues de la transformation de la tyrosine sous l’action de 3 enzymes clés : la tyrosinase, Tyrp1 (pour Tyrosinase-related protein 1), et Dct/Tyrp2 (pour DOPAchrome tautomérase/Tyrosinase-related protein 2) (voir« synthèse des mélanines »).

Au cours de leur synthèse dans les mélanocytes , les mélanines sont stockées dans les mélanosomes . Une fois maturés, ces derniers sont transportés le long des microtubules et du cytosquelette d’actine afin d’être stockés au niveau d’expansions cytoplasmiques appelées « dendrites mélanocytaires ». Le transport sur les fibres d’actine se fait grâce à un complexe moléculaire associant au minimum un moteur moléculaire, la myosine Va, une petite GTPase, Rab27a, et la mélanophiline. Les mélanosomes sont ensuite transférés aux kératinocytes des couches supérieures de l’épiderme via un processus qui reste encore mal connu. Plusieurs hypothèses ont été formulées : 1) cytophagocytose : le kératinocyte phagocyte l’extrémité dendritique mélanocytaire contenant les mélanosomes ; 2) fusion membranaire : les membranes cellulaires des mélanocytes et des kératinocytes fusionnent engendrant la formation d’un canal cytoplasmique qui permet le passage des mélanosomes ; 3) ingestion : les mélanosomes sont déversés sous forme de vésicules qui sont phagocytées par ces derniers ; 4) injection directe : les mélanosomes sont transférés via un processus d’exocytose-endocytose couplées. Au sein des kératinocytes, essentiellement basaux, en se rassemblant au niveau apical du noyau (phénomène de capping), les mélanines constituent un véritable filtre pour les rayonnements UV. Elles sont également capables d’absorber les radicaux libres générés dans les cellules par les radiations UV afin d’empêcher les lésions de l’ADN. Sous l’effet des UV, une réponse adaptative se met en place avec une stimulation de la mélanogenèse. En effet, au sein des mélanocytes, une irradiation UV induit une augmentation de l’expression du facteur de transcription MITF capable d’activer la transcription des gènes impliqués dans la mélanogenèse tels que la tyrosinase, Tyrp1 et Dct/Tyrp-2. Les kératinocytes répondent également à une exposition aux UV en augmentant la synthèse des hormones alpha-MSH (alpha melanocyte stimulating hormone) et ACTH (adrénocorticotrope), puissants activateurs de la mélanogenèse, ainsi que l’expression de leur récepteur MC1R à la surface des mélanocytes.
V. Une barrière antimicrobienne
L’épiderme est sans cesse exposé aux micro-organismes dont certains peuvent être pathogènes. Pour lutter contre cette agression, les cellules épidermiques impliquées dans l’immunité innée, à savoir les kératinocytes et les cellules de Langerhans, jouent un rôle majeur de barrière immunologique. Tout comme les kératinocytes, les cellules de Langerhans, véritables sentinelles immunologiques, reconnaissent des structures moléculaires particulières à la surface des micro-organismes par des récepteurs de danger (protéines PRR pour « Pattern Recognition Receptors »), en particulier les récepteurs de type TLR (Toll-Like receptor) ou de type NLR (Nod-Like Receptors). Ces cellules, une fois activées, sont capables de phagocytose et de présentation des antigènes microbiens aux lymphocytes (cellules de Langerhans), produisent des peptides antimicrobiens (kératinocytes), des chimiokines (les deux types) ou d’autres médiateurs chimiques.
Cette composante immunologique est complétée par une composante structurale : le processus de desquamation qui assure l’élimination des cornéocytes de surface et des micro-organismes installés, et la structure quasi-infranchissable que représente la couche cornée. En plus de cette barrière physique, le pH acide et le faible taux d’humidité de la couche cornée, et la température de la peau inférieure à 37°C sont défavorables à la croissance bactérienne. Finalement, la présence de peptides et lipides antimicrobiens font de l’épiderme un véritable défenseur contre l’infection.
V.1. Le pH de la couche cornée
La couche cornée présente, en surface, un pH acide de 4,5-5. Ceci permet de maintenir la symbiose du microbiote cutané en favorisant le développement de la microflore commensale tout en inhibant la croissance des bactéries pathogènes, notamment du staphylocoque doré préférant un environnement neutre. Le pH joue également un rôle dans la régulation des protéases impliquées dans la desquamation. Le pH neutre des couches profondes de la couche cornée assure une forte interaction entre les kallikréines 5 et 7 et leur inhibiteur LEKTI. En revanche, en remontant vers la surface, l’acidification réduit cette interaction « protéases-inhibiteurs ». Ceci permet la libération des kallikréines et l’activation du processus de desquamation assurant l’élimination des micro-organismes de surface.
V.2. Les peptides antimicrobiens
Les peptides antimicrobiens, concentrés au niveau de la couche cornée, jouent un rôle majeur dans l’immunité innée. Ces petits peptides amphiphiles sont composés d’acides aminés cationiques et hydrophobes présentant un arrangement spatial bien défini. Produits par les kératinocytes, ainsi que les sébocytes et les cellules des glandes sudorales, ils présentent une activité antimicrobienne avec un spectre d’action très large comprenant les bactéries, les champignons et les virus. La plupart détruisent les organismes cibles en perturbant leur intégrité membranaire notamment grâce à une interaction électrostatique avec les lipides. Il s’agit d’un mécanisme de défense dynamique. En effet, bien que certains peptides antimicrobiens soient exprimés de manière constitutive, beaucoup d’entre eux présentent une expression inductible. A ce jour, plus d’une centaine de peptides antimicrobiens ont été identifiés au niveau cutané. Les principaux sont les défensines et les cathélicidines mais il existe d’autres molécules présentant des propriétés antimicrobiennes évidentes. On retrouve notamment des inhibiteurs de protéases tels que l’élafine, SKALP (skin-derived antileukoprotease) ou encore SLPI (secretory leucocyte protease inhibitor), des chimiokines telles que la psoriasine (S100A7), des ribonucléases comme la RNase 7, du lysozyme, etc (#Glaser et al, 2005).
Chez l’Homme, les défensines les plus exprimées dans la peau sont les β-défensines, petits peptides de 3 à 6 kDa chargés positivement et composés de trois feuillets β repliés par des ponts disulfures. Les kératinocytes synthétisent notamment hBD-1, -2 et -3. Alors que hBD-1 est constitutivement exprimée, hBD-2 et hBD-3 sont induites sous l’effet de stimuli pro-inflammatoires (IL1, TNF-α, INF-γ, lipopolysaccharides) et sécrétées par l’intermédiaire des corps lamellaires. hBD-2 présente un spectre d’action beaucoup plus restreint que hBD-3. En effet, alors que hBD-3 est active contre les bactéries à Gram positif ou négatif et contre les champignons, hBD-2 présente une activité antimicrobienne essentiellement dirigée contre les bactéries à Gram négatif.
Les cathélicidines sont des peptides linéaires en hélice α. Dans l’épiderme humain, seule la cathélicidine hCAP-18 (human cathelicidin antimicrobial protein 18kDa) a été identifiée. Elle est exprimée de manière constitutive au niveau des follicules pilo-sébacés et inductible dans l’épiderme interfolliculaire. Cette induction dépend de conditions inflammatoires impliquant IGF-1, TGF-α et la Vitamine D3. hCAP-18 est sécrétée à l’interface de la couche granuleuse et de la couche cornée par l’intermédiaire des corps lamellaires. Elle est ensuite protéolysée par les kallikréines et les protéases des neutrophiles comme la protéinase 3 afin de donner naissance à un peptide carboxy-terminal LL37 qui possède un large spectre d‘action sur les bactéries à Gram positif ou à Gram négatif.
Plusieurs études convergent pour indiquer un rôle crucial des peptides antimicrobiens épidermiques in vivo. L’implication de LL37 dans la protection vis-à-vis des micro-organismes pathogènes a été démontrée dans un modèle murin où l’absence d’expression de cette protéine entraîne une augmentation de la sensibilité des souris à l’infection par Streptococcus pyogenes. Alors que les infections cutanées sont plutôt rares chez les patients atteints de psoriasis qui présentent une expression forte de peptides antimicrobiens, les patients atteints de dermatite atopique aigüe qui montrent une moindre induction de hBD-2, hBD-3 et LL37, sont sensibles aux infections cutanées, notamment à Staphylococcus aureus (#Schröder, 2011). En outre, des analyses in vitro montrent que l’activité antimicrobienne de LL37 est renforcée en présence des défensines, suggérant une synergie entre les différents peptides antimicrobiens afin de former une barrière antimicrobienne efficace.
En plus de leur activité antimicrobienne, ces peptides antimicrobiens cutanés possèdent d’autres fonctions biologiques. Ils sont capables de moduler l’inflammation par la chimio-attraction de cellules immunitaires et la régulation de la production des cytokines inflammatoires, de réguler la prolifération cellulaire, la cicatrisation, la ré-épithélialisation et la néovascularisation . Les peptides antimicrobiens sont ainsi considérés comme de véritables molécules multifonctionnelles.
V.3. Les chimiokines
Les chimiokines constituent une superfamille de cytokines chimiotactiques produites après stimulation par des signaux de dangers. Elles régulent l’activation, la différenciation ainsi que la migration des cellules immunitaires (cellules phagocytaires, cellules T, éosinophiles). Il existe 2 classes majeures de chimiokines en fonction de la distance séparant deux résidus cystéine conservés : les chimiokines α qui contiennent une séquence CXC (cystéine-résidu aléatoire-cystéine) et les chimiokines β qui ont une séquence CC (cystéine-cystéine).
Au sein de l’épiderme, la présence de micro-organismes pathogènes, tel que le staphylocoque doré, induit notamment une augmentation de l’expression de CXCL8 (ou IL8) par les kératinocytes. CXCL8 agit de manière paracrine pour assurer le recrutement des polynucléaires neutrophiles au niveau du site de l’infection. Outre ce rôle dans le recrutement des cellules immunitaires, certaines chimiokines telles que CCL20, CXCL7, CXCL9-11 et CCL20, possèdent un effet antimicrobien avec un large spectre d‘action sur les bactéries à Gram positif ou à Gram négatif.
V.4. Les lipides épidermiques
Outre leur rôle dans la mise en place d’une barrière hydrique, les lipides épidermiques sont également impliqués dans la régulation de l’immunité innée. En effet, les acides gras libres ainsi que les sphingosines présentent une véritable activité antimicrobienne. Des expériences in vitro ont mis en évidence l’efficacité des acides gras libres, notamment de l’acide palmitoléique, contre les bactéries à Gram positif et des sphingosines contre ces mêmes bactéries et contre les bactéries à Gram négatif. De plus, in vivo, la sensibilité accrue aux infections cutanées, notamment à Staphylococcus aureus, des patients atteints de dermatite atopique, est associée à une diminution du niveau de sphingosine.
Conclusion
La raison d’être de l’épiderme est, comme nous venons de le voir, d’assurer un rôle de barrière multifonctionnelle grâce aux cellules immunitaires et aux kératinocytes qui le composent. La mise en place de cette barrière et son fonctionnement nécessitent des mécanismes physiques, chimiques et biochimiques complexes et finement régulés. Ses altérations, cause ou conséquence de nombreuses maladies de la peau très rares comme certaines génodermatoses ou très fréquentes comme le psoriasis et l’eczéma, sont la preuve de son importance (#Le Lamer et al, 2015 ; #Pendaries et al, 2014).
Bibliographie
Bouwstra JA, Gooris GS, Dubbelaar FE, Ponec M. Phase behavior of lipid mixtures based on human ceramides : coexistence of crystalline and liquid phases. J Lipid Res. 2001 Nov ;42(11):1759-70. PubMed PMID : 11714845.
Brown SJ, McLean WH. One remarkable molecule : filaggrin. J Invest Dermatol. 2012 Mar ;132(3 Pt 2):751-62. doi : 10.1038/jid.2011.393. Epub 2011 Dec 8. Review. PubMed PMID : 22158554 ; PubMed Central PMCID : PMC3378480.
Candi E, Schmidt R, Melino G. The cornified envelope : a model of cell death in the skin. Nat Rev Mol Cell Biol. 2005 Apr ;6(4):328-40. doi:10.1038/nrm1619. PMID:15803139
Feingold KR, Elias PM. Role of lipids in the formation and maintenance of the cutaneous permeability barrier. Biochim Biophys Acta. 2014 Mar ;1841(3):280-94. doi : 10.1016/j.bbalip.2013.11.007. Epub 2013 Nov 18. Review. PubMed PMID : 24262790.
Furuse M, Hata M, Furuse K, Yoshida Y, Haratake A, Sugitani Y, Noda T, Kubo A, Tsukita S. Claudin-based tight junctions are crucial for the mammalian epidermal barrier : a lesson from claudin-1-deficient mice. J Cell Biol. 2002 Mar 18 ;156(6):1099-111. Epub 2002 Mar 11. PubMed PMID : 11889141 ; PubMed CentralPMCID : PMC2173463.
Gläser R, Harder J, Lange H, Bartels J, Christophers E, Schröder JM. Antimicrobial psoriasin (S100A7) protects human skin from Escherichia coli infection. Nat Immunol. 2005 Jan ;6(1):57-64. Epub 2004 Nov 28. PubMed PMID :
15568027.
Kirschner N, Brandner JM. Barriers and more : functions of tight junction
proteins in the skin. Ann N Y Acad Sci. 2012 Jun ;1257:158-66. doi :
10.1111/j.1749-6632.2012.06554.x. Review. PubMed PMID : 22671602.
Le Lamer M, Pellerin L, Reynier M, Cau L, Pendaries V, Leprince C, Méchin MC,
Serre G, Paul C, Simon M. Defects of corneocyte structural proteins and epidermal
barrier in atopic dermatitis. Biol Chem. 2015 Nov ;396(11):1163-79. doi :
10.1515/hsz-2015-0141. PubMed PMID : 26020560.
Madison KC. Barrier function of the skin : « la raison d’être » of the epidermis. J Invest Dermatol. 2003 Aug ;121(2):231-41. Review. PubMed PMID : 12880413.
Pendaries V, Malaisse J, Pellerin L, Le Lamer M, Nachat R, Kezic S, Schmitt
AM, Paul C, Poumay Y, Serre G, Simon M. Knockdown of filaggrin in a
three-dimensional reconstructed human epidermis impairs keratinocyte
differentiation. J Invest Dermatol. 2014 Dec ;134(12):2938-46. doi :
10.1038/jid.2014.259. Epub 2014 Jun 13. PubMed PMID : 24940654.
Rawlings AV, Matts PJ. Stratum corneum moisturization at the molecular level : an update in relation to the dry skin cycle. J Invest Dermatol. 2005 Jun ;124(6):1099-110. Review. PubMed PMID : 15955083.
[Schröder JM. Antimicrobial peptides in healthy skin and atopic dermatitis.Allergol Int. 2011 Mar ;60(1):17-24. doi : 10.2332/allergolint.10-RAI-0292. Epub 2011 Jan 25. Review. PubMed PMID : 21252615.
Steinert PM, Marekov LN. The proteins elafin, filaggrin, keratin intermediate filaments, loricrin, and small proline-rich proteins 1 and 2 are isodipeptidecross-linked components of the human epidermal cornified cell envelope. J Biol Chem. 1995 Jul 28 ;270(30):17702-11. PubMed PMID : 7543090.
van Smeden J, Janssens M, Gooris GS, Bouwstra JA. The important role of stratum corneum lipids for the cutaneous barrier function. Biochim Biophys Acta. 2014 Mar ;1841(3):295-313. doi : 10.1016/j.bbalip.2013.11.006. Epub 2013 Nov 16. Review. PubMed PMID : 24252189.
Vermeij WP, Alia A, Backendorf C. ROS quenching potential of the epidermal cornified cell envelope. J Invest Dermatol. 2011 Jul ;131(7):1435-41. doi :
10.1038/jid.2010.433. Epub 2011 Jan 20. PubMed PMID : 21248766.
Mots-clés
- filaggrine
- lipides intercornéocytaires
- desmosome/desmosomes
- acide urocanique
- céramide/céramides
- cornéodesmosine
- cornéodesmosome
- couche cornée / stratum corneum
- enveloppe cornée
- épiderme
- espèces réactives de l’oxygène/dérivés réactifs de l’oxygène/DROs/ROS
- Facteur Naturel d’Hydratation
- involucrine
- kératine/kératines
- kératinocyte/kératinocytes
- la transglutaminase/les transglutaminases
- loricrine
- mélanine/mélanines
- mélanocyte/mélanocytes
- mélanosome/mélanosomes
- peau humaine
- pigmentation cutanée/pigmentation
- rayonnement ultraviolet /UV/UVA/UVB/UVC
- stress oxydant/stress oxydatif
- système anti-oxydant/systèmes anti-oxydants
![]() fr
fr
![]() STRUCTURE ET FONCTIONS
STRUCTURE ET FONCTIONS
![]() Fonctions de la peau
?
Fonctions de la peau
?
Site réalisé avec SPIP 3.0.17 + AHUNTSIC

Visiteurs connectés : 33