Les carcinomes basocellulaires
Les carcinomes basocellulaires (CBC) sont des tumeurs d’origine kératinocytaire. Ils représentent 75% des cancers épidermiques d’origine non mélanocytaire (Green and Battistutta, 1990 ; Miller, 1991a ; Miller, 1991b ; Preston and Stern, 1992). C’est le cancer le plus fréquent chez l’Homme et son incidence est en constante augmentation (+10% depuis les années 1960). Aux Etats-Unis, environ 500000 nouveaux cas de CBC sont recensés chaque année (Diepgen and Mahler, 2002 ; Wong et al., 2003). Cependant, il reste difficile d’évaluer de façon précise la fréquence des CBC car elle varie selon les régions du globe et parce qu’ils ne sont pas toujours mentionnés dans les registres sur les cancers. Le rapport d’incidence homme/femme est de 1,9 aux Etats-Unis,1,2 au Royaume-Uni et 1,7 en Australie (Buettner and Raasch, 1998 ; Holme et al.,2000 ; Diepgen and Mahler, 2002 ; Rubin et al., 2005).
1. Généralités
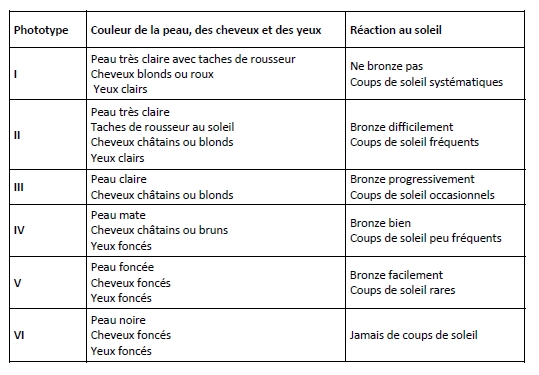
L’étiologie des CBC est liée au phototype , à l’âge et à l’exposition aux UV (Miller, 1995 ; DePinho, 2000). Concernant le phototype, Kimonis et al. ont montré dans une étude comparative que les individus d’origine caucasienne (phototypes I à IV) présentaient un risque plus important de développer des CBC que ceux d’origine afro‐américaine (phototypes V et VI) (Kimonis et al., 1997).
Les CBC ne se développent que très rarement avant 50 ans mais leur incidence augmente fortement avec l’âge (Buettner and Raasch, 1998 ; Diepgen and Mahler, 2002). Les UV jouant un rôle majeur dans l’étiologie des CBC, 90% de ces tumeurs se développent dans des zones photo‐exposées notamment au niveau du crâne, du visage, du cou, du tronc, parfois sur les bras et les jambes mais rarement sur le dos ou les mains (Giles et al., 1988 ; Holme et al., 2000 ; Rubin et al., 2005).
Pour les CBC, il n’y a pas de lésion précurseur connue. Dans la plupart des cas, les kératinocytes de CBC ressemblent aux kératinocytes de la couche basale (noyau volumineux et cytoplasme réduit), ils franchissent la membrane basale et prolifèrent dans le derme en formant des amas (nodules) ou des travées. L’organisation palissadique des kératinocytes situés à la périphérie des zones invaginées et le décollement dermo‐épidermique sont deux caractéristiques histologiques des CBC.
Il existe plusieurs types de CBC selon leur histologie et leurs propriétés d’invasion : superficiel, nodulaire, micro‐nodulaire, morphéiforme, infiltrant et métatypique (pour illustration voir les figures de Wong et al., 2003 ;Crowson, 2006. ;Dermato-Basocellulaire-Wikinu). Le potentiel invasif fait de ces trois dernières formes les plus agressives et les plus graves.
Ce sont des tumeurs à croissance lente et qui métastasent rarement (0,003 à 0,55% des cas) mais pouvant être localement très invasives dans le derme sous‐jacent et s’avérer très délabrantes (Robinson and Dahiya, 2003) notamment lorsqu’ils sont situés au niveau du visage.
2. Le rôle des ultraviolets dans le développement de carcinomes basocellulaires
Le principal facteur de risque de développement de cancers cutanés d’origine non mélanocytaire (CCNM) est l’exposition aux ultraviolets (UV) solaires. En effet, les altérations moléculaires, cellulaires et tissulaires induites par ces radiations contribuent, à long terme, au développement de ces cancers.
Les UV ne représentent qu’une faible partie du rayonnement solaire arrivant à la surface de la Terre constitué à 63% de lumière visible et 32% d’infrarouges (Sage, 1993). On distingue les UVA (320‐400 nm), les UVB (280‐320 nm) et les UVC (100‐280 nm). Les UVC ainsi qu’une partie des UVB sont absorbés par la couche d’ozone de la stratosphère. Environ 95% d’UVA et 5% d’UVB atteignent la surface du globe mais ils ne représentent respectivement que 5 et 0,3% du rayonnement solaire nous parvenant.
La peau constitue la première barrière contre les UV, les couches de l’épiderme sont capables d’absorber une partie des rayonnements (< 300 nm) mais laissent passer les moins énergétiques (Young et al., 1998). Les UV sont à l’origine de lésions dans l’ADN, majoritairement des dimères cyclobutaniques de pyrimidines (CPD, cyclobutane pyrimidine dimer) et de pyrimidines (6‐4)‐pyrimidone (6‐4‐PP) (pour revue voir Warrick, 2012), réparées principalement par le mécanisme de réparation par excision de nucléotides (NER, nucleotide excision repair) ; pour revue voir Cleaver et al., 2009 . Ils peuvent également générer des espèces réactives de l’oxygène (ROS, reactive oxygen species) entraînant la formation de lésions appelées 8‐oxo‐7,8‐dihydroguanine (8‐oxoG) et réparées par un mécanisme de réparation par excision de bases (BER, base excision repair) (pour revue voir Simon, 2008 ; Hoeijmakers, 2001). Si les lésions CPD et 6-4-PP ne sont pas réparées avant la réplication, elles peuvent conduire à des mutations dans l’ADN (transition C—>T ou mutation en tandem CC—>TT, signatures des UV).
Les dommages à l’ADN induits par les UV conduisent à des dérégulations au niveau de la voie de signalisation impliquant la protéine p53 dite « gardien du génome ». p53 est impliquée dans le contrôle du cycle cellulaire (arrêt en phases G1/S et G2/M), la réparation de l’ADN et l’apoptose en cas d’excès de lésions ; pour revue voir : (Murray‐Zmijewski et al., 2008). TP53 est un gène suppresseur de tumeur muté dans 33 à 56% des CBC sporadiques, dans la plupart des cas sur des séquences spécifiques appelées « points chauds » (Rady et al., 1992 ; Ziegler et al., 1993 ; Zhang et al.,
2001 ; Kim et al., 2002 ; Bolshakov et al., 2003 ; Reifenberger et al., 2005).
Au cours des dernières décennies, l’incidence des CBC n’a cessé d’augmenter du fait de l’allongement de la durée de la vie mais également en rapport avec des changements d’habitudes au sein de la population tels que l’utilisation de cabines de bronzage ainsi que des modifications de style vestimentaire. Par ailleurs, dans les populations à peau claire, l’incidence des CCNM augmente avec la proximité de l’équateur. Ainsi, en Australie, la diminution de 8° de latitude s’accompagne d’un accroissement de 2,4 du risque de développer un CCNM (Giles et al., 1988).
Les CBC se développent majoritairement après 50 ans du fait de l’accumulation de lésions induites par les UV au cours de la vie et leur incidence augmente lors du vieillissement . Ainsi, une personne âgée de plus de 75 ans présentera un risque de développer un CBC 4,5 fois plus élevé qu’une personne de 50 ans (Holme et al., 2000). De plus, les personnes ayant déjà eu un CBC ont un risque accru d’en développer un autre.
3. La dépendance stromale des carcinomes basocellulaires
En 1959, constatant le faible potentiel métastatique des CBC, Pinkus introduit la notion dedépendance stromale des cellules tumorales (Pinkus, 1959). Cette hypothèse sera renforcée dans les années suivantes par des expériences d’autogreffes (Lyles et al., 1960 ; Van Scott and Reinertson, 1961). Lyles et al., ont prélevé, sur 18 patients volontaires, des CBC qu’ils leur ont regreffé immédiatement dans une zone saine en sous‐cutané ou en intradermique. Les CBC transplantés avec leur stroma se sont maintenus alors que ceux greffés sans la partie stromale ont régressé. De plus, contrairement aux kératinocytes de CSC, ceux isolés à partir de CBC ne sont pas cultivables in vitro. Cela suggère, qu’en dehors de leur environnement tumoral, les kératinocytes de CBC sont incapables
de survivre (Lyles et al., 1960).
Le stroma est un tissu conjonctif et vasculaire qui joue un rôle de soutien et de nutrition indispensable à la croissance tumorale. Ce microenvironnement est constitué de trois éléments essentiels qui sont :
- la matrice extracellulaire et les fibroblastes sécrétant diverses molécules dont des facteurs de croissance,
- les vaisseaux néoformés (angiogénèse tumorale) indispensables pour l’apport de nutriments et d’oxygène aux cellules cancéreuses,
- les cellules inflammatoires et immunocompétentes responsables de la réaction stromale lors de l’invasion du tissu conjonctif sain par les cellules tumorales.
Ces dernières années, plusieurs équipes ont démontré le rôle essentiel du stroma dans le développement et la prolifération tumorale suggérant des échanges de signaux très intenses entre les cellules cancéreuses et celles du microenvironnement selon un modèle de régulation paracrine (Gaggioli et al., 2007 ; Gaggioli, 2008 ; Yauch et al., 2008 ; Tian et al., 2009) ; pour revues voir : (Kalluri and Zeisberg, 2006 ; Rubin and de Sauvage, 2006).
4. Les pathologies associées à une prédisposition au carcinome basocellulaire
Il existe plusieurs génodermatoses qui prédisposent au développement des CBC :
4.1 La naevomatose basocellulaire
La naevomatose basocellulaire (NBC), ou syndrome de Gorlin, est une maladie héréditaire à transmission autosomique dominante associée à des mutations dans le gène suppresseur de tumeur PTCH1 (« gatekeeper ») impliqué dans la voie de signalisation Sonic Hedgehog/PATCHED (SHH/PTCH) (Gorlin, 1995 ; Hahn et al., 1996). Les patients présentent une forte prédisposition aux CBC mais pas au CSC.
4.2 Le xeroderma pigmentosum
Le xeroderma pigmentosum (XP) est un syndrome génétique rare à transmission autosomique récessive associé à des mutations des gènes « carekeepers » impliqués dans le principal mécanisme de réparation des lésions de l’ADN UV‐induites, la réparation par excision de nucléotides (NER). Les patients qui s’exposent au soleil développent de nombreux cancers cutanés (CBC, CSC, mélanomes) souvent avant l’âge de 8 ans et leur pronostic vital est réduit (de Boer and Hoeijmakers, 2000).
4.3 Le syndrome de Bazex‐Dupré‐Christol
Le syndrome de Bazex‐Dupré‐Christol est une génodermatose rare à transmission dominante liée au chromosome X qui prédispose au développement précoce de multiples CBC. Moins de 150 cas ont été décrits et le gène impliqué dans cette maladie n’a pas encore été identifié avec certitude (Vabres et al., 1995 ; Castori et al., 2009).
4.4 L’albinisme
L’albinisme regroupe un ensemble de maladies héréditaires à transmission autosomique récessive. L’albinisme est caractérisé par l’hypo‐pigmentation voir l’absence totale de pigmentation de la peau, des phanères et des yeux. Les patients sont prédisposés au développement de tumeurs cutanées UV‐induites de façon précoce (CBC, CSC et mélanomes).
4.5 Le syndrome de Brooke‐Spiegler
Le syndrome de Brooke‐Spiegler est une génodermatose rare à transmission autosomique dominante associée à des mutations dans le gène CYLD. La protéine CYLD est impliqué dans plusieurs voies de signalisation, notamment les voies NF‐κB et MAPK (immunité et réponse au stress cellulaire). Les patients présentent une forte prédisposition aux trichoépithéliomes éventuellement accompagnés d’autres tumeurs annexielles (van den Ouweland et al., 2011). Ils peuvent également développer des CBC.
4.6 Les patients à multiples CBC
Il existe des patients développant de multiples CBC, ≥ 10 CBC (Heitzer et al., 2007), pour lesquels aucune mutation dans un gène impliqué dans une génodermatose n’a pu être identifiée et qui ne présentent aucun autre symptôme clinique. Il est difficile d’estimer le nombre de cas car ils sont très peu étudiés dans la littérature. Certains cas sont dus à des mutations (n’affectant pas PTCH1) de novo et d’autres semblent être héréditaires.
5. Les traitements des carcinomes basocellulaires
5.1 L’exérèse chirurgicale
L’exérèse chirurgicale est la méthode la plus utilisée pour enlever un carcinome basocellulaire. Il existe deux types d’opérations : l’exérèse classique et la chirurgie micrographique de Mohs. Cette dernière donne d’excellents résultats en termes de récidive car elle repose sur l’analyse immédiate de la tumeur par microscopie afin d’en déterminer les bords et d’enlever la totalité du tissu cancéreux. Le pourcentage de récidive d’un CBC n’est que de 1 à 3% avec cette méthode contre 10% avec l’exérèse classique (van der Geer et al., 2009). La chirurgie micrographique de Mohs permet de préserver le tissu sain tout en éradiquant la tumeur offrant ainsi d’excellents résultats au niveau sanitaire et esthétique.
5.2 Les lasers
Les lasers présentent plusieurs avantages par rapport à la chirurgie : ils ne nécessitent pas d’anesthésie, ne laissent pas de cicatrice et un seul passage permet d’éliminer plusieurs lésions superficielles et ce en une seule séance (Szeimies and Karrer, 2006 ; Amini et al., 2010). Moskalik et al. ont traité 3461 patients présentant des lésions faciales (3534 CBC au total) et ont obtenu un taux de récidive de seulement 2% (Moskalik et al., 2009).
5.3 La photothérapie dynamique
Tout comme les lasers, la photothérapie dynamique permet le traitement de CBC superficiels. Elle repose sur l’utilisation d’une molécule photo‐active telle que l’acide 5‐δ aminolévulinique (ALA) et d’une source d’irradiation lumineuse (635 nm pour l’ALA). La molécule est appliquée en topique sur la zone à traiter ou injectée en intraveineuse puis la zone à traiter est irradiée, activant ainsi le composé photo‐actif qui va générer des espèces réactives de l’oxygène qui vont conduire à la destruction des cellules cancéreuses (Szeimies and Karrer, 2006 ; Lien and Sondak, 2011).
5.4 Les autres traitements
Certaines techniques telles que la radiothérapie ou la cryochirurgie sont de moins en moins utilisées car leur efficacité et le taux de récidive (8% et 18% respectivement) sont plus élevés qu’avec les méthodes vues précédemment (Avril et al., 1997). Elle est pratiquée dans des cas de CBC de grande taille ou localisés dans des zones inopérables.
La méthode d’électrocoagulation‐curetage est également abandonnée au profit de techniques plus récentes car elle ne peut s’appliquer que dans le cas de petites tumeurs (< 2 cm), que le taux de récidive est de 13% et qu’elle peut laisser des cicatrices.
Les traitements par des molécules chimiques en application topique (5‐fluorouracil (Efudix®), imiquimod (AldaraTM)) ou en injections intra‐ ou péri‐lésionelle (interférons) sont parfois employés mais le taux de récidive, leur réelle spécificité et les effets secondaires à long terme ne sont pas encore connus (Amini et al., 2010). Les rétinoïdes (vitamine A acide et ses dérivés de synthèse) sont utilisés en application topique dans les traitements préventifs et curatifs des CBC notamment chez les patients atteints de naevomatose basocellulaire. Parmi ces composés, le plus efficace est le tazarotène (Zorac®) qui cible de façon spécifique les récepteurs de l’acide rétinoïque (RAR, retinoic acid receptors) de type β/γ. Le tazarotène provoque, in vitro et in vivo, l’inhibition de la prolifération des cellules de CBC et l’induction de leur apoptose (Orlandi et al., 2004 ; So et al., 2008).
Bibliographie
DePinho, R.A. 2000. The age of cancer. Nature. 408:248‐54.
Diepgen, T.L., and V. Mahler. 2002. The epidemiology of skin cancer. Br J Dermatol. 146 Suppl 61:1‐6.
Gorlin, R.J. 1995. Nevoid basal cell carcinoma syndrome. Dermatol Clin. 13:113‐25.
Hoeijmakers, J.H. 2001. Genome maintenance mechanisms for preventing cancer. Nature. 411:366‐
74.
Kalluri, R., and M. Zeisberg. 2006. Fibroblasts in cancer. Nat Rev Cancer. 6:392‐401.
Miller, S.J. 1991a. Biology of basal cell carcinoma (Part I). J Am Acad Dermatol. 24:1‐13.
Miller, S.J. 1991b. Biology of basal cell carcinoma (Part II). J Am Acad Dermatol. 24:161‐75.
Miller, S.J. 1995. Etiology and pathogenesis of basal cell carcinoma. Clin Dermatol. 13:527‐36.
[Pinkus, H. 1959. Clinical, histologic and differential considerations, Washington D.C.]
Preston, D.S., and R.S. Stern. 1992. Nonmelanoma cancers of the skin. N Engl J Med. 327:1649‐62.
Rubin, A.I., E.H. Chen, and D. Ratner. 2005. Basal‐cell carcinoma. N Engl J Med. 353:2262‐9.
Simon M. 2008. Réparation de l’ADN
Warrick E (2012) : Effets des UV sur l’ADN : lésions et mutations
Wong CS, Strange RC, Lear JT (2003). Basal cell carcinoma. Bmj 327 : 794-8.
Portfolio
![]() fr
fr
![]() LES PRINCIPALES MALADIES DE LA PEAU
LES PRINCIPALES MALADIES DE LA PEAU
![]() Les cancers cutanés
?
Les cancers cutanés
?
Site réalisé avec SPIP 3.0.17 + AHUNTSIC

Visiteurs connectés : 26